

Arrest of 5HT neuron differentiation delays respiratory maturation and impairs neonatal homeostatic responses to environmental challenges
- PMID: 17656160
- PMCID: PMC2593840
- DOI: 10.1016/j.resp.2007.06.002
Arrest of 5HT neuron differentiation delays respiratory maturation and impairs neonatal homeostatic responses to environmental challenges
Abstract
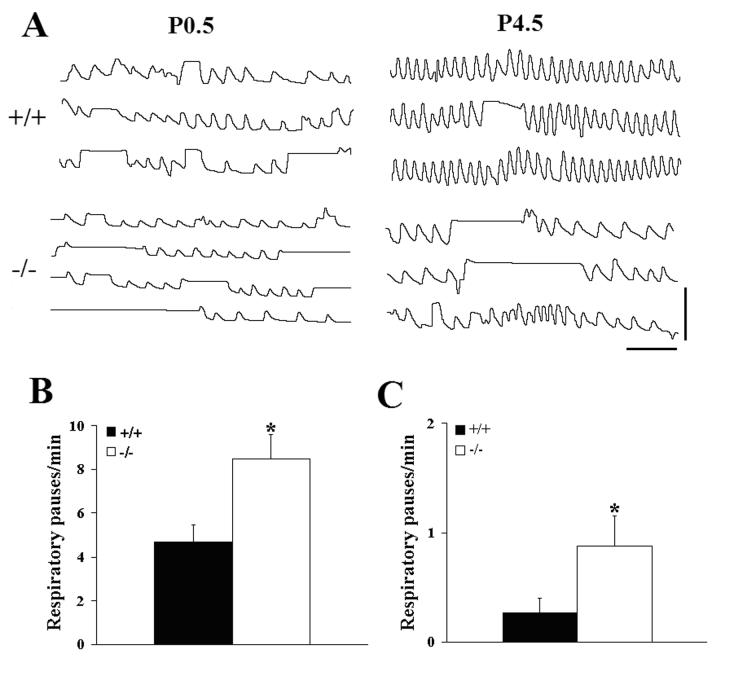
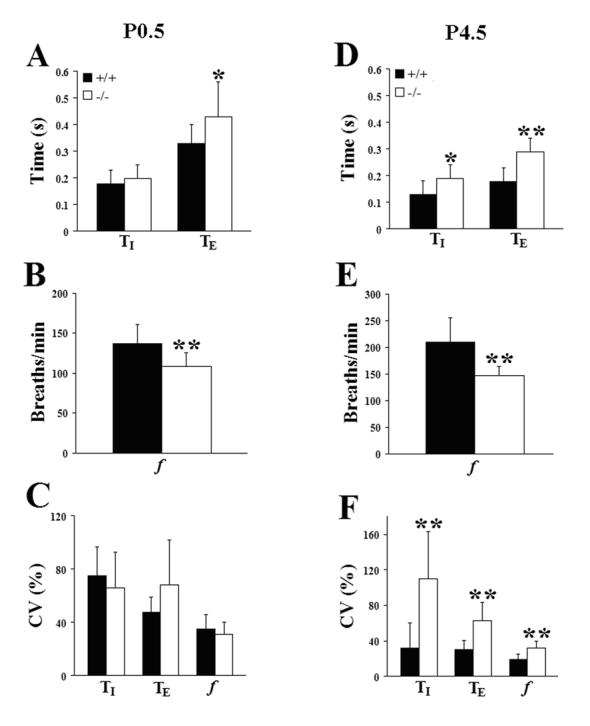
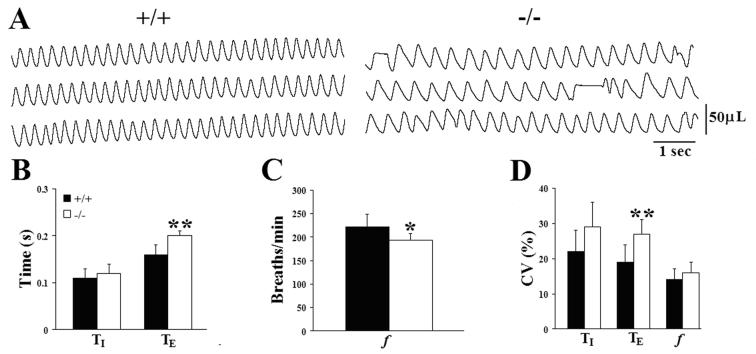
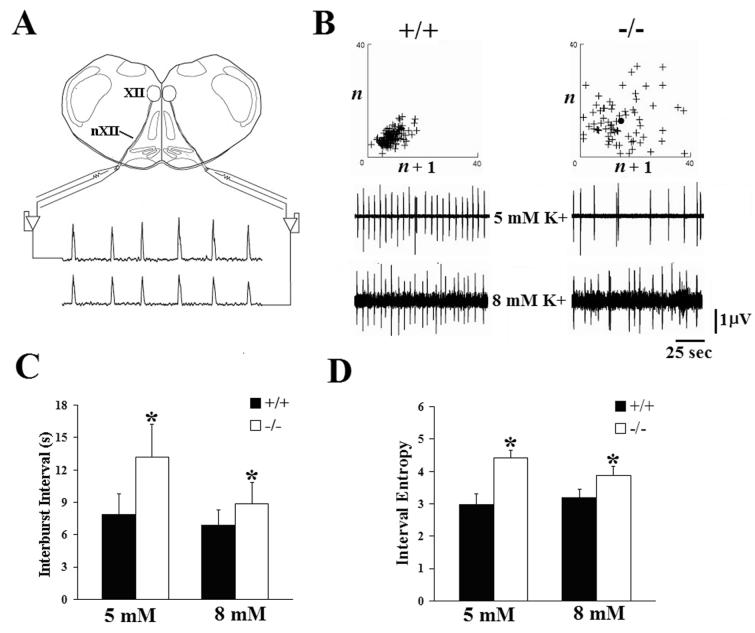
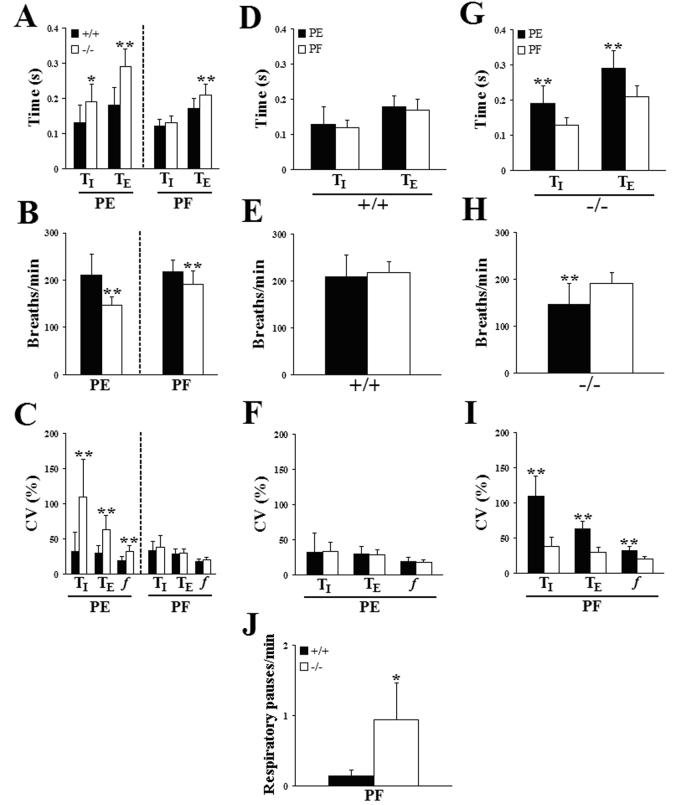
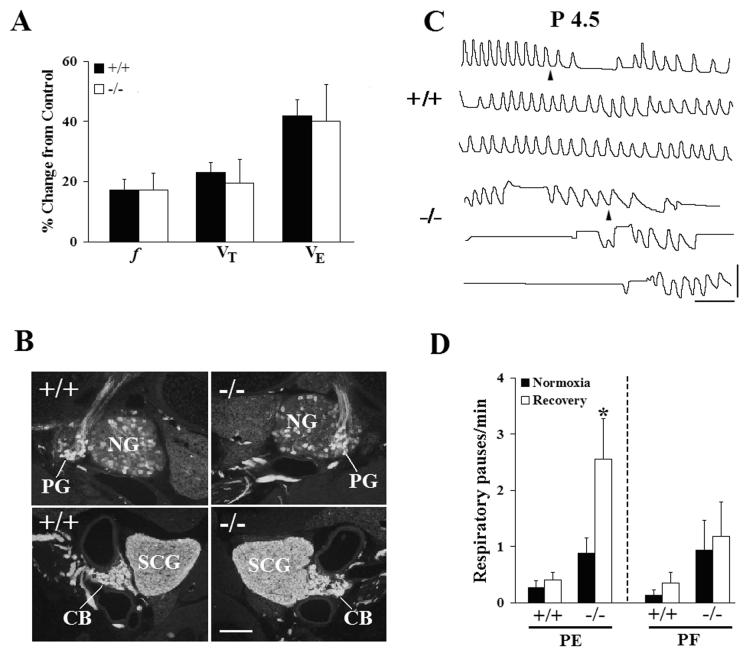
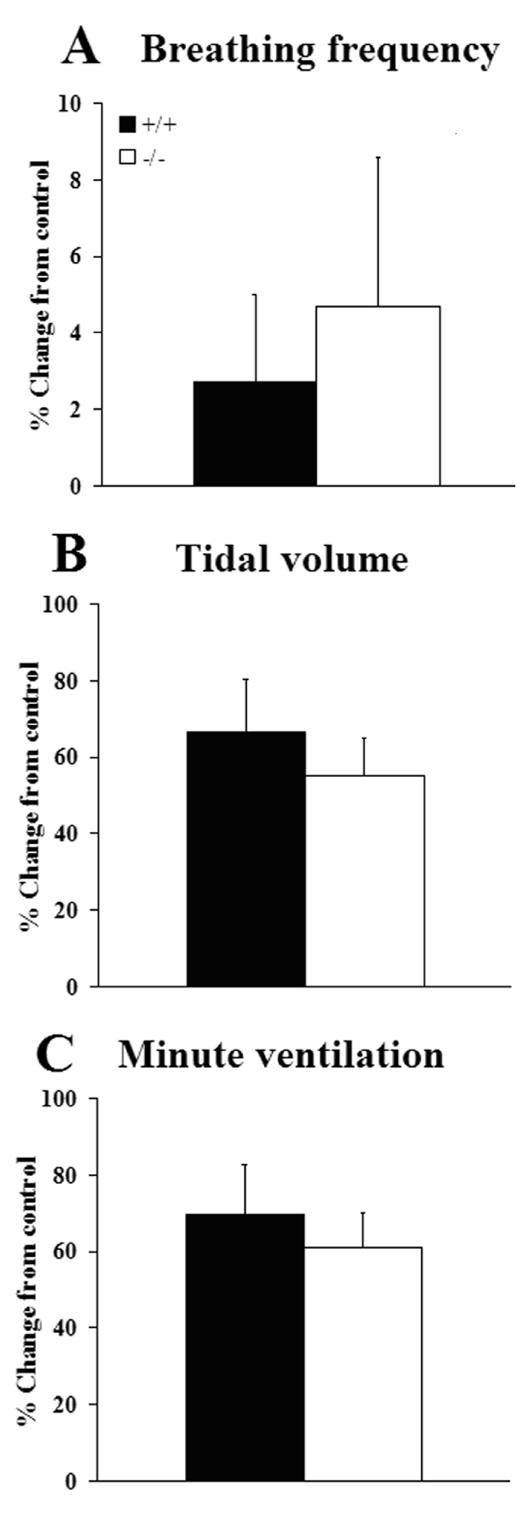
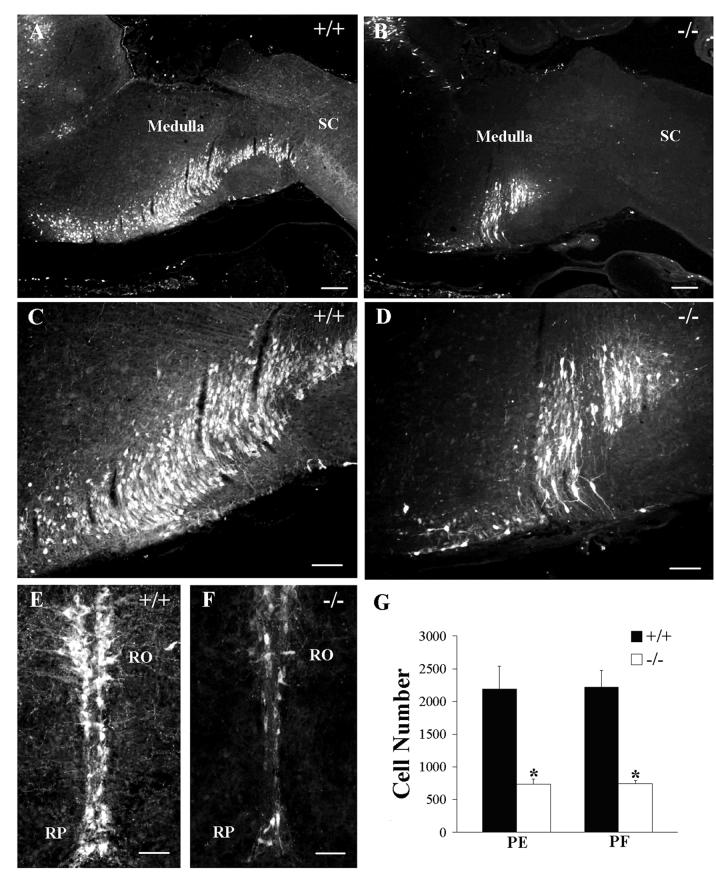
Serotonin (5HT) is a powerful modulator of respiratory circuitry in vitro but its role in the development of breathing behavior in vivo is poorly understood. Here we show, using 5HT neuron-deficient Pet-1 (Pet-1(-/-)) neonates, that serotonergic function is required for the normal timing of postnatal respiratory maturation. Plethysmographic recordings reveal that Pet-1(-/-) mice are born with a depressed breathing frequency and a higher incidence of spontaneous and prolonged respiratory pauses relative to wild type littermates. The wild type breathing pattern stabilizes by postnatal day 4.5, while breathing remains depressed, highly irregular and interrupted more frequently by respiratory pauses in Pet-1(-/-) mice. Analysis of in vitro hypoglossal nerve discharge indicates that instabilities in the central respiratory rhythm generator contribute to the abnormal Pet-1(-/-) breathing behavior. In addition, the breathing pattern in Pet-1(-/-) neonates is susceptible to environmental conditions, and can be further destabilized by brief exposure to hypoxia. By postnatal day 9.5, however, breathing frequency in Pet-1(-/-) animals is only slightly depressed compared to wild type, and prolonged respiratory pauses are rare, indicating that the abnormalities seen earlier in the Pet-1(-/-) mice are transient. Our findings provide unexpected insight into the development of breathing behavior by demonstrating that defects in 5HT neuron development can extend and exacerbate the period of breathing instability that occurs immediately after birth during which respiratory homeostasis is vulnerable to environmental challenges.
Figures
References
-
- Abadie V, Champagnat J, Fortin G. Branchiomotor activities in mouse embryo. NeuroReport. 2000;11:141–145. - PubMed
-
- Al-Zubaidy Z, Erickson R, Greer J. Serotonergic and noradrenergic effects on respiratory neural discharges in the medullary slice preparation of neonatal rats. Pflügers Arch. 1996;431:942–949. - PubMed
-
- Bartlett D, Remmers JE, Jr., Gautier H. Laryngeal regulation of respiratory airflow. Respir. Physiol. 1973;18:194–200. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
