

Mechanisms of neuronal chloride accumulation in intact mouse olfactory epithelium
- PMID: 17656441
- PMCID: PMC2277205
- DOI: 10.1113/jphysiol.2007.129601
Mechanisms of neuronal chloride accumulation in intact mouse olfactory epithelium
Abstract
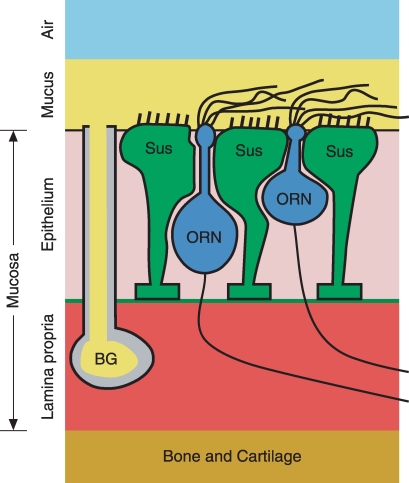
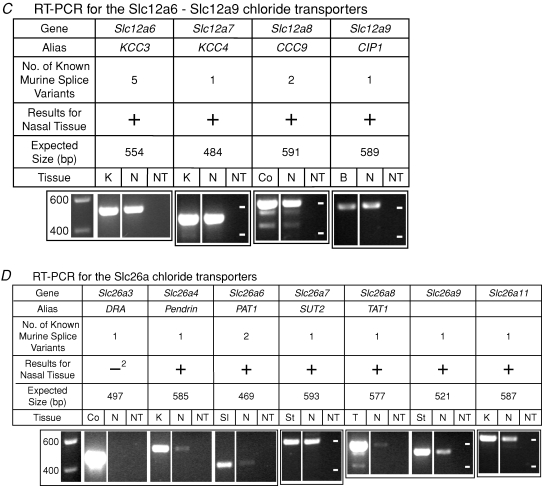
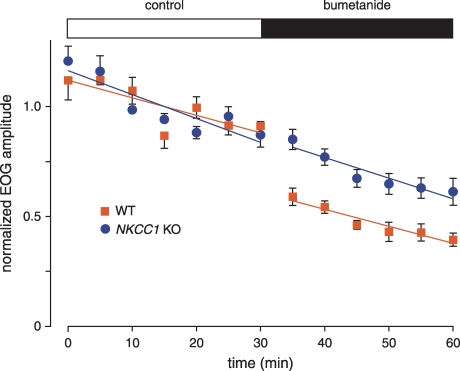
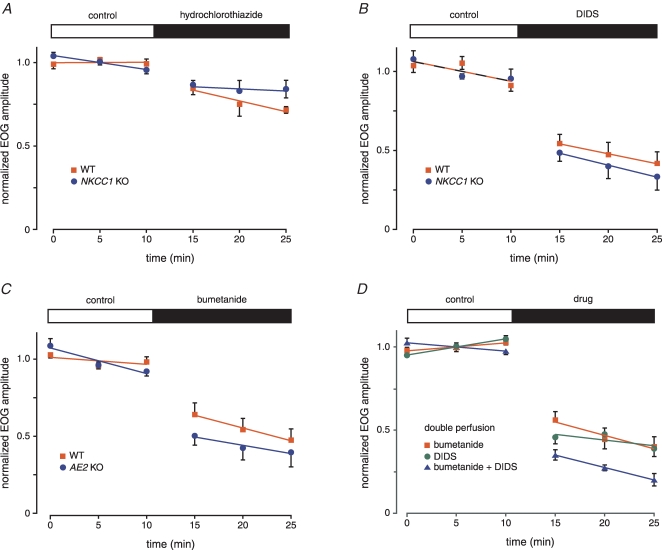
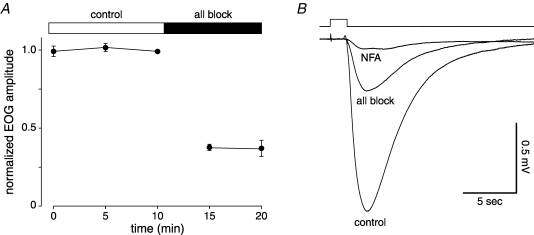
When olfactory receptor neurons respond to odours, a depolarizing Cl(-) efflux is a substantial part of the response. This requires that the resting neuron accumulate Cl(-) against an electrochemical gradient. In isolated olfactory receptor neurons, the Na(+)-K(+)-2Cl(-) cotransporter NKCC1 is essential for Cl(-) accumulation. However, in intact epithelium, a robust electrical olfactory response persists in mice lacking NKCC1. This response is largely due to a neuronal Cl(-) efflux. It thus appears that NKCC1 is an important part of a more complex system of Cl(-) accumulation. To identify the remaining transport proteins, we first screened by RT-PCR for 21 Cl(-) transporters in mouse nasal tissue containing olfactory mucosa. For most of the Cl(-) transporters, the presence of mRNA was demonstrated. We also investigated the effects of pharmacological block or genetic ablation of Cl(-) transporters on the olfactory field potential, the electroolfactogram (EOG). Mice lacking the common Cl(-)/HCO(3)(-) exchanger AE2 had normal EOGs. Block of NKCC cotransport with bumetanide reduced the EOG in epithelia from wild-type mice but had no effect in mice lacking NKCC1. Hydrochlorothiazide, a blocker of the Na(+)-Cl(-) cotransporter, had only a small effect. DIDS, a blocker of some KCC cotransporters and Cl(-)/HCO(3)(-) exchangers, reduced the EOG in epithelia from both wild-type and NKCC1 knockout mice. A combination of bumetanide and DIDS decreased the response more than either drug alone. However, no combination of drugs completely abolished the Cl(-) component of the response. These results support the involvement of both NKCC1 and one or more DIDS-sensitive transporters in Cl(-) accumulation in olfactory receptor neurons.
Figures

Similar articles
-
Mice lacking NKCC1 have normal olfactory sensitivity.Physiol Behav. 2008 Jan 28;93(1-2):44-9. doi: 10.1016/j.physbeh.2007.07.011. Epub 2007 Aug 1. Physiol Behav. 2008. PMID: 17719611 Free PMC article.
-
AE2 Cl-/HCO3- exchanger is required for normal cAMP-stimulated anion secretion in murine proximal colon.Am J Physiol Gastrointest Liver Physiol. 2010 Apr;298(4):G493-503. doi: 10.1152/ajpgi.00178.2009. Epub 2010 Jan 28. Am J Physiol Gastrointest Liver Physiol. 2010. PMID: 20110461 Free PMC article.
-
Mechanism of the excitatory Cl- response in mouse olfactory receptor neurons.Neuron. 2005 Feb 17;45(4):553-61. doi: 10.1016/j.neuron.2005.01.012. Neuron. 2005. PMID: 15721241 Free PMC article.
-
The bumetanide-sensitive Na-K-2Cl cotransporter NKCC1 as a potential target of a novel mechanism-based treatment strategy for neonatal seizures.Neurosurg Focus. 2008 Sep;25(3):E22. doi: 10.3171/FOC/2008/25/9/E22. Neurosurg Focus. 2008. PMID: 18759624 Review.
-
The ins and outs of intracellular chloride in olfactory receptor neurons.Neuron. 2005 Feb 17;45(4):481-2. doi: 10.1016/j.neuron.2005.02.002. Neuron. 2005. PMID: 15721233 Review.
Cited by
-
Deep sequencing of the murine olfactory receptor neuron transcriptome.PLoS One. 2015 Jan 15;10(1):e0113170. doi: 10.1371/journal.pone.0113170. eCollection 2015. PLoS One. 2015. PMID: 25590618 Free PMC article.
-
Standardized, systemic phenotypic analysis of Slc12a1I299F mutant mice.J Biomed Sci. 2014 Aug 2;21(1):68. doi: 10.1186/s12929-014-0068-0. J Biomed Sci. 2014. PMID: 25084970 Free PMC article.
-
Mice lacking NKCC1 have normal olfactory sensitivity.Physiol Behav. 2008 Jan 28;93(1-2):44-9. doi: 10.1016/j.physbeh.2007.07.011. Epub 2007 Aug 1. Physiol Behav. 2008. PMID: 17719611 Free PMC article.
-
ANO2 is the cilial calcium-activated chloride channel that may mediate olfactory amplification.Proc Natl Acad Sci U S A. 2009 Jul 14;106(28):11776-81. doi: 10.1073/pnas.0903304106. Epub 2009 Jun 26. Proc Natl Acad Sci U S A. 2009. PMID: 19561302 Free PMC article.
-
Differential expression of pancreatic protein and chemosensing receptor mRNAs in NKCC1-null intestine.World J Gastrointest Pathophysiol. 2016 Feb 15;7(1):138-49. doi: 10.4291/wjgp.v7.i1.138. World J Gastrointest Pathophysiol. 2016. PMID: 26909237 Free PMC article.
References
-
- Alper SL, Darman RB, Chernova MN, Dahl NK. The AE family of Cl−/HCO3− exchangers. J Nephrol. 2002;15(Suppl. 5):S41–S53. - PubMed
-
- Bakalyar HA, Reed RR. Identification of a specialized adenylyl cyclase that may mediate odorant detection. Science. 1990;250:1403–1406. - PubMed
-
- Boccaccio A, Menini A. Temporal development of CNG and Ca2+-activated Cl− currents in isolated mouse olfactory sensory neurons. J Neurophysiol. 2007;98:153–160. - PubMed
-
- Brunet LJ, Gold GH, Ngai J. General anosmia caused by a targeted disruption of the mouse olfactory cyclic nucleotide-gated cation channel. Neuron. 1996;17:681–693. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases