

Complex signatures of selection and gene conversion in the duplicated globin genes of house mice
- PMID: 17660536
- PMCID: PMC2013706
- DOI: 10.1534/genetics.107.078550
Complex signatures of selection and gene conversion in the duplicated globin genes of house mice
Abstract
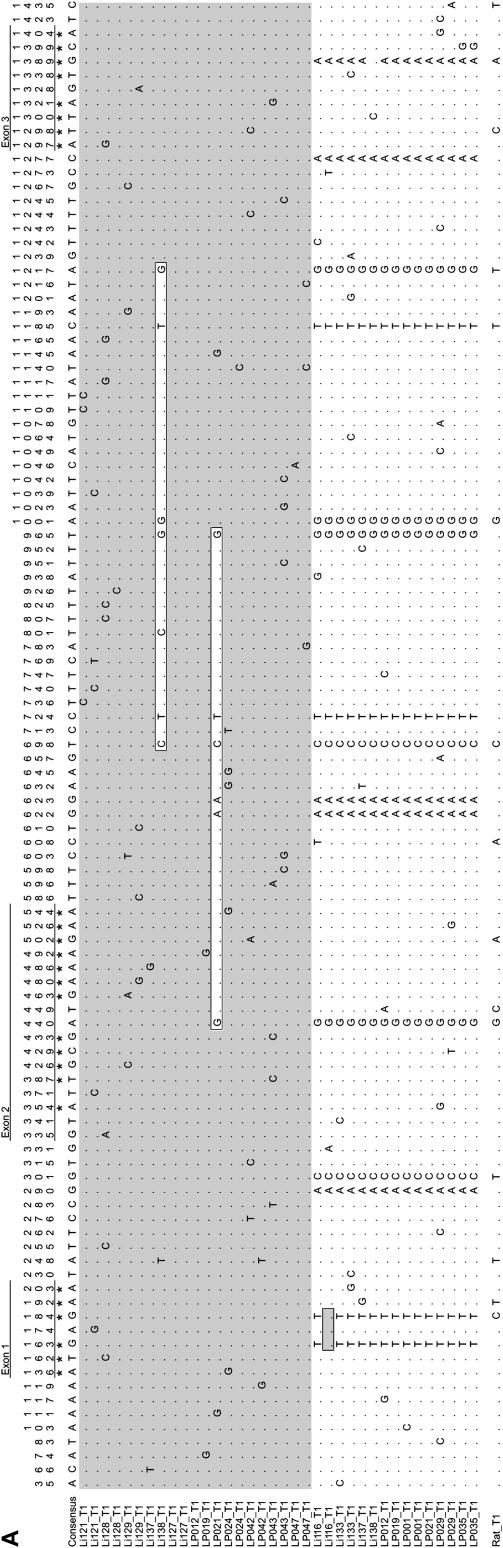
Results of electrophoretic surveys have suggested that hemoglobin polymorphism may be maintained by balancing selection in natural populations of house mice, Mus musculus. Here we report a survey of nucleotide variation in the adult globin genes of house mice from South America. We surveyed nucleotide polymorphism in two closely linked alpha-globin paralogs and two closely linked beta-globin paralogs to test whether patterns of variation are consistent with a model of long-term balancing selection. Surprisingly high levels of nucleotide polymorphism at the two beta-globin paralogs were attributable to the segregation of two highly divergent haplotypes, Hbbs (which carries two identical beta-globin paralogs) and Hbbd (which carries two functionally divergent beta-globin paralogs). Interparalog gene conversion on the Hbbs haplotype has produced a highly unusual situation in which the two paralogs are more similar to one another than either one is to its allelic counterpart on the Hbbd haplotype. Levels of nucleotide polymorphism and linkage disequilibrium at the two beta-globin paralogs suggest a complex history of diversity-enhancing selection that may be responsible for long-term maintenance of alternative protein alleles. The alternative two-locus beta-globin haplotypes are associated with pronounced differences in intraerythrocyte glutathione and nitric oxide metabolism, suggesting a possible mechanism for selection on hemoglobin function.
Figures









References
-
- Aguileta, G., J. P. Bielawski and Z. H. Yang, 2006. Proposed standard nomenclature for the alpha- and beta-globin gene families. Genes Genet. Syst. 81: 367–371. - PubMed
-
- Auffray, J. C., F. Vanlerberghe and J. Brittondavidian, 1990. The house mouse progression in Eurasia: a paleontological and archaeozoological approach. Biol. J. Linn. Soc. 41: 13–25.
Publication types
MeSH terms
Substances
Associated data
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
