

Sodium channel Nav1.6 accumulates at the site of infraorbital nerve injury
- PMID: 17662136
- PMCID: PMC1941742
- DOI: 10.1186/1471-2202-8-56
Sodium channel Nav1.6 accumulates at the site of infraorbital nerve injury
Abstract
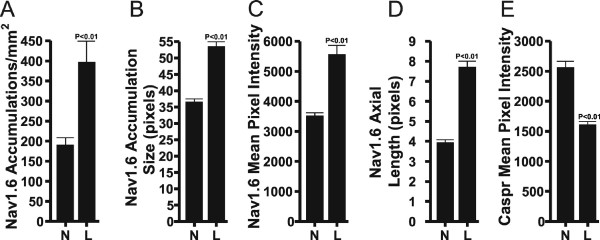
Background: Sodium channel (NaCh) expressions change following nerve and inflammatory lesions and this change may contribute to the activation of pain pathways. In a previous study we found a dramatic increase in the size and density of NaCh accumulations, and a remodeling of NaChs at intact and altered myelinated sites at a location just proximal to a combined partial axotomy and chromic suture lesion of the rat infraorbital nerve (ION) with the use of an antibody that identifies all NaCh isoforms. Here we evaluate the contribution of the major nodal NaCh isoform, Nav1.6, to this remodeling of NaChs following the same lesion. Sections of the ION from normal and ION lesioned subjects were double-stained with antibodies against Nav1.6 and caspr (contactin-associated protein; a paranodal protein to identify nodes of Ranvier) and then z-series of optically sectioned images were captured with a confocal microscope. ImageJ (NIH) software was used to quantify the average size and density of Nav1.6 accumulations, while additional single fiber analyses measured the axial length of the nodal gap, and the immunofluorescence intensity of Nav1.6 in nodes and of caspr in the paranodal region.
Results: The findings showed a significant increase in the average size and density of Nav1.6 accumulations in lesioned IONs when compared to normal IONs. The results of the single fiber analyses in caspr-identified typical nodes showed an increased axial length of the nodal gap, an increased immunofluorescence intensity of nodal Nav1.6 and a decreased immunofluorescence intensity of paranodal caspr in lesioned IONs when compared to normal IONs. In the lesioned IONs, Nav1.6 accumulations were also seen in association with altered caspr-relationships, such as heminodes.
Conclusion: The results of the present study identify Nav1.6 as one isoform involved in the augmentation and remodeling of NaChs at nodal sites following a combined partial axotomy and chromic suture ION lesion. The augmentation of Nav1.6 may result from an alteration in axon-Schwann cell signaling mechanisms as suggested by changes in caspr expression. The changes identified in this study suggest that the participation of Nav1.6 should be considered when examining changes in the excitability of myelinated axons in neuropathic pain models.
Figures


References
-
- Tzoumaka E, Tischler AC, Sangameswaran L, Eglen RM, Hunter JC, Novakovic SD. Differential distribution of the tetrodotoxin-sensitive rPN4/NaCh6/Scn8a sodium channel in the nervous system. J Neurosci Res. 2000;60:37–44. doi: 10.1002/(SICI)1097-4547(20000401)60:1<37::AID-JNR4>3.0.CO;2-W. - DOI - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Medical
Miscellaneous
