

Divergent functions and distinct localization of the Notch ligands DLL1 and DLL3 in vivo
- PMID: 17664336
- PMCID: PMC2064846
- DOI: 10.1083/jcb.200702009
Divergent functions and distinct localization of the Notch ligands DLL1 and DLL3 in vivo
Abstract
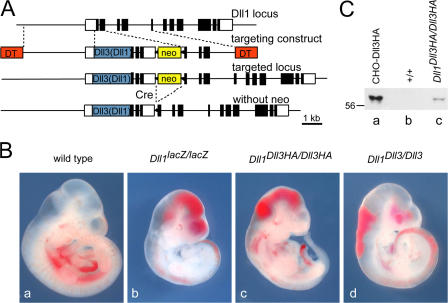
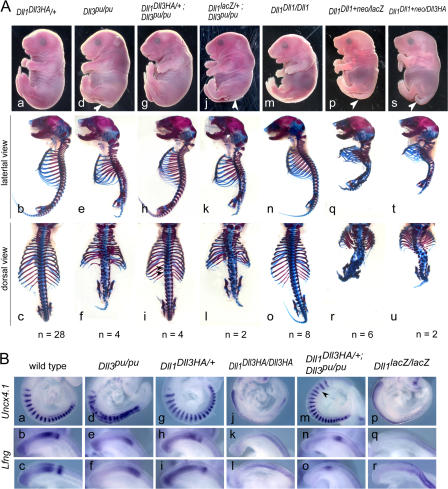
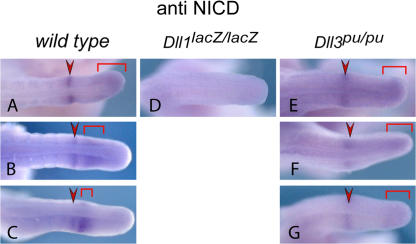
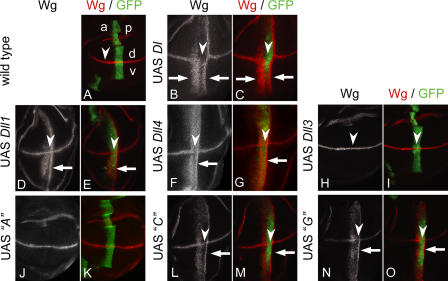
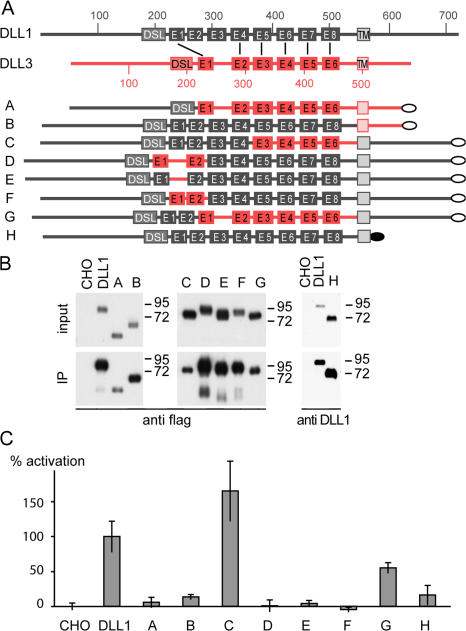
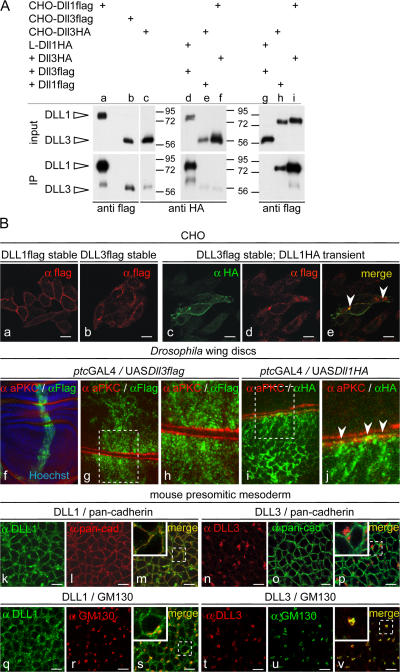
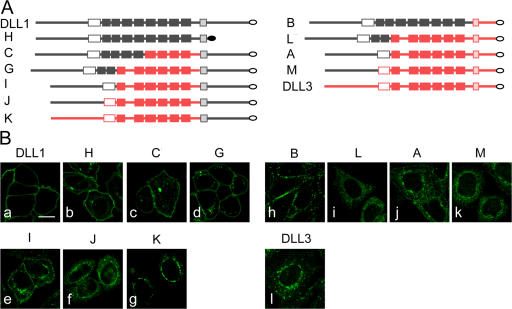
The Notch ligands Dll1 and Dll3 are coexpressed in the presomitic mesoderm of mouse embryos. Despite their coexpression, mutations in Dll1 and Dll3 cause strikingly different defects. To determine if there is any functional equivalence, we replaced Dll1 with Dll3 in mice. Dll3 does not compensate for Dll1; DLL1 activates Notch in Drosophila wing discs, but DLL3 does not. We do not observe evidence for antagonism between DLL1 and DLL3, or repression of Notch activity in mice or Drosophila. In vitro analyses show that differences in various domains of DLL1 and DLL3 individually contribute to their biochemical nonequivalence. In contrast to endogenous DLL1 located on the surface of presomitic mesoderm cells, we find endogenous DLL3 predominantly in the Golgi apparatus. Our data demonstrate distinct in vivo functions for DLL1 and DLL3. They suggest that DLL3 does not antagonize DLL1 in the presomitic mesoderm and warrant further analyses of potential physiological functions of DLL3 in the Golgi network.
Figures
References
-
- Abdelkhalek, H.B., A. Beckers, K. Schuster-Gossler, M.N. Pavlova, H. Burkhardt, H. Lickert, J. Rossant, R. Reinhardt, L.C. Schalkwyk, I. Müller, et al. 2004. The mouse homeobox gene Not is required for caudal notochord development and affected by the truncate mutation. Genes Dev. 18:1725–1736. - PMC - PubMed
-
- Artavanis-Tsakonas, S., M.D. Rand, and R.J. Lake. 1999. Notch signaling: cell fate control and signal integration in development. Science. 284:770–776. - PubMed
-
- Ascano, J.M., L.J. Beverly, and A.J. Capobianco. 2003. The C-terminal PDZ- ligand of JAGGED1 is essential for cellular transformation. J. Biol. Chem. 278:8771–8779. - PubMed
-
- Barsi, J.C., R. Rajendra, J.I. Wu, and K. Artzt. 2005. Mind bomb1 is a ubiquitin ligase essential for mouse embryonic development and Notch signaling. Mech. Dev. 122:1106–1117. - PubMed
-
- Blaumueller, C.M., and S. Artavanis-Tsakonas. 1997. Comparative aspects of Notch signaling in lower and higher eukaryotes. Perspect. Dev. Neurobiol. 4:325–343. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
