

Advanced glycation endproducts induce podocyte apoptosis by activation of the FOXO4 transcription factor
- PMID: 17667983
- PMCID: PMC3191877
- DOI: 10.1038/sj.ki.5002456
Advanced glycation endproducts induce podocyte apoptosis by activation of the FOXO4 transcription factor
Abstract
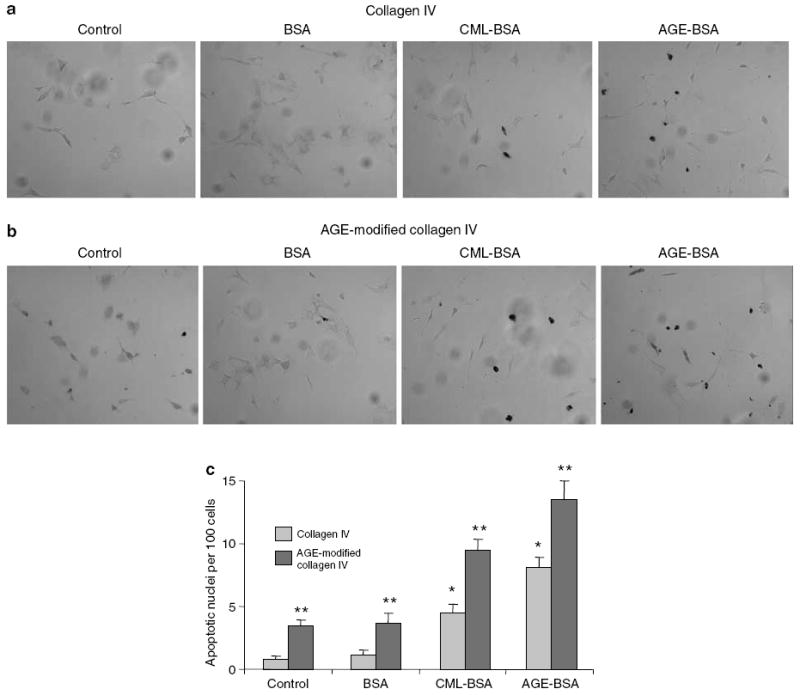
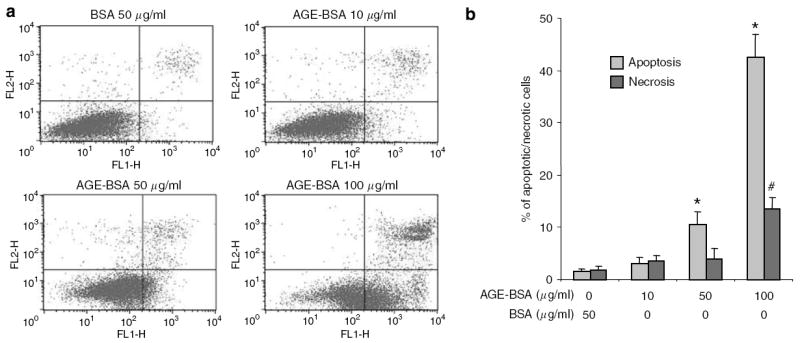
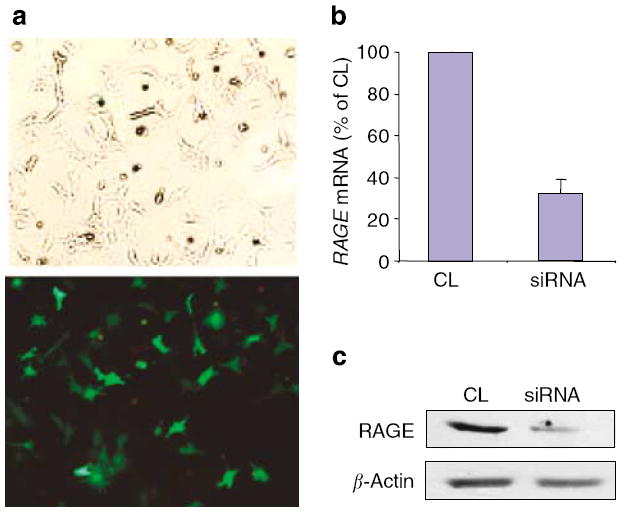
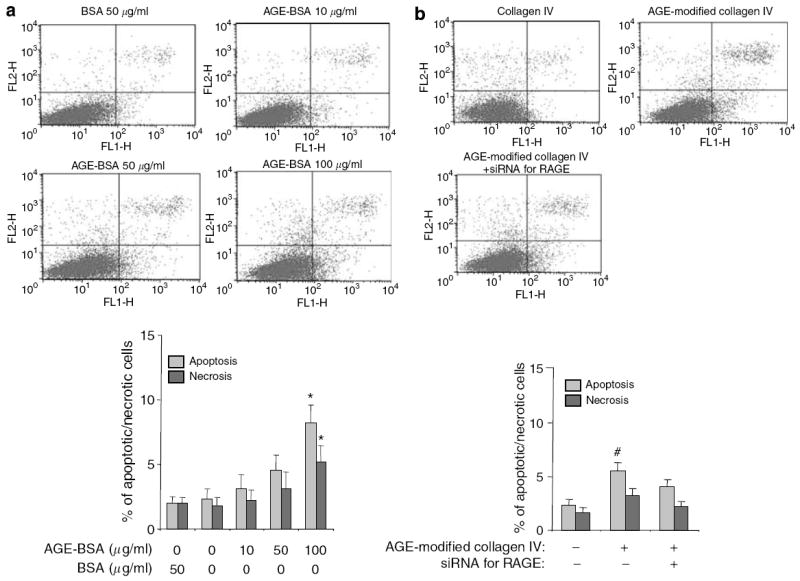
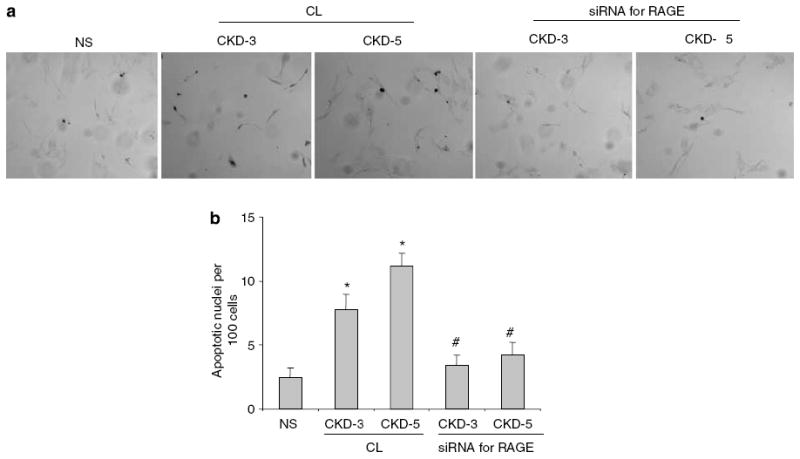
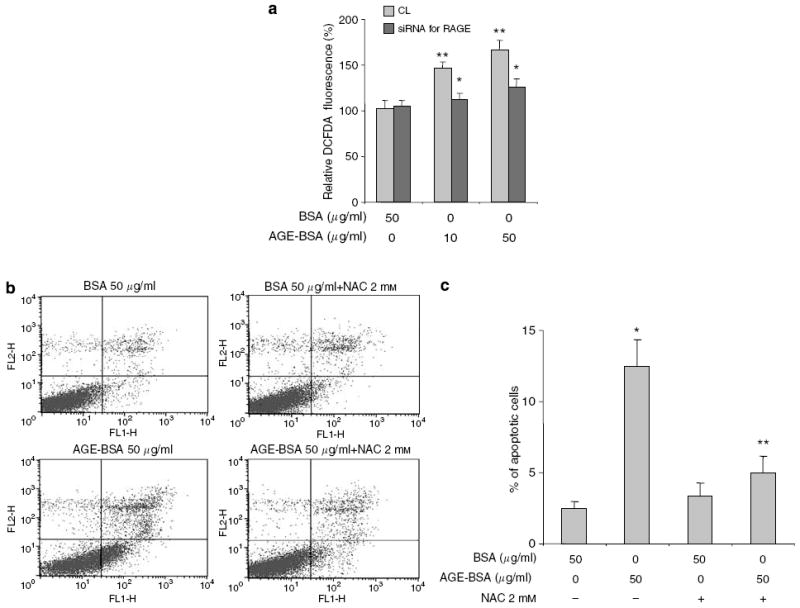
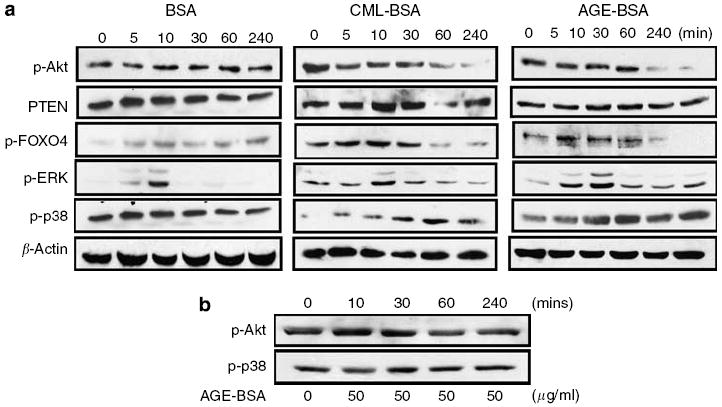
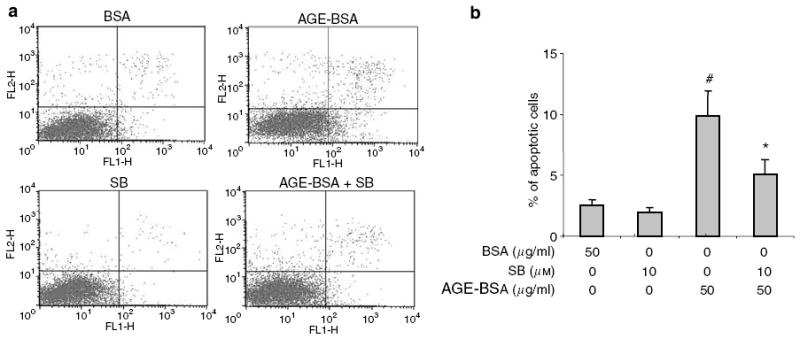
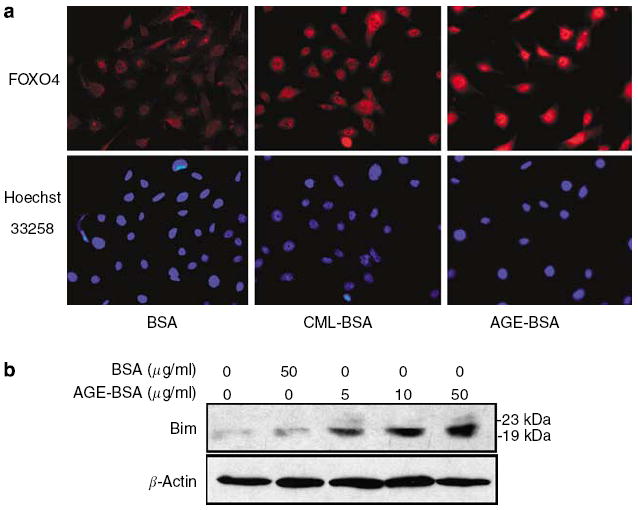
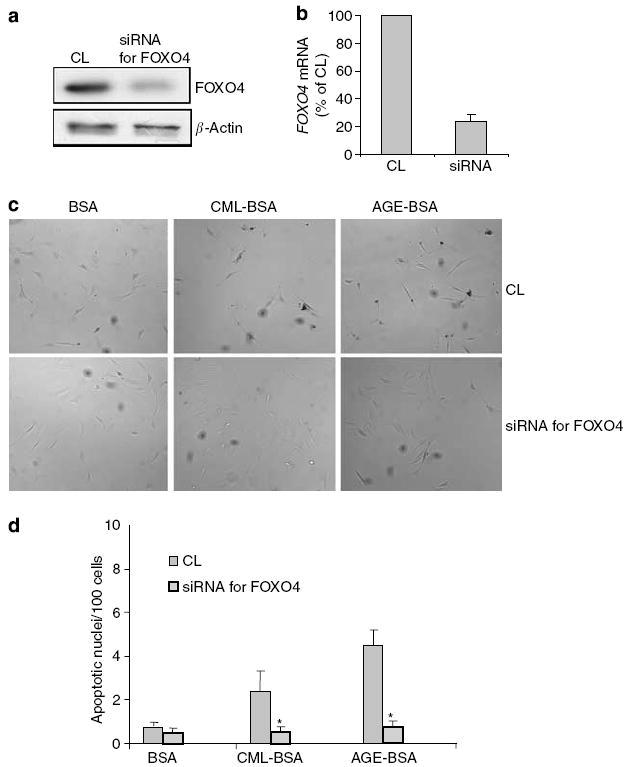
Advanced glycation endproducts (AGEs) and a receptor for AGEs (RAGE) have been linked in the pathogenesis of diabetic nephropathy. RAGE is usually localized to podocytes and is increased in diabetes. RAGE activation increases reactive oxygen species production, which mediates hyperglycemia-induced podocyte apoptosis in early diabetic nephropathy. Here, we examined the interaction of AGE and RAGE on podocyte apoptosis. When we exposed murine cultured podocytes to bovine serum albumin (BSA) that was modified by AGEs or to carboxymethyl-lysine BSA, more apoptosis was found when compared with unmodified BSA. Similarly, more podocytes underwent detachment and apoptosis when cultured on AGE-modified collagen IV than on native collagen IV. AGEs isolated from sera of patients with chronic kidney disease also caused apoptosis of podocytes. Apoptosis was diminished by small interference RNA (siRNA) for RAGE in podocytes exposed to AGE-BSA, but not to AGE-modified collagen IV. Both AGE- and carboxymethyl-lysine modified-BSA activated p38MAP kinase and inhibition of this kinase reduced the apoptotic effect of AGE-BSA. Exposure to AGE-BSA was associated with Akt dephosphorylation and FOXO4 transcriptional activation leading to an increase in the expression of an effector protein of apoptosis, Bim. siRNA for FOXO4 abolished AGE-BSA-induced apoptosis of podocytes. Our study suggests that an AGE-RAGE interaction contributes to podocyte apoptosis by activation of the FOXO4 transcription factor.
Figures
References
-
- USRDS: the United States Renal Data System. Am J Kidney Dis. 2003;43:1–230. - PubMed
-
- Ziyadeh FN. Mediators of diabetic renal disease: the case for TGF-beta as the major mediator. J Am Soc Nephrol. 2004;15:S55–S57. - PubMed
-
- Drummond K, Mauer M. the early natural history of nephropathy in type 1 diabetes. II. Early renal structural changes in type 1 diabetes. Diabetes. 2002;51:1580–1587. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
