

Capture of linear fragments at a double-strand break in yeast
- PMID: 17670800
- PMCID: PMC1976456
- DOI: 10.1093/nar/gkm521
Capture of linear fragments at a double-strand break in yeast
Abstract
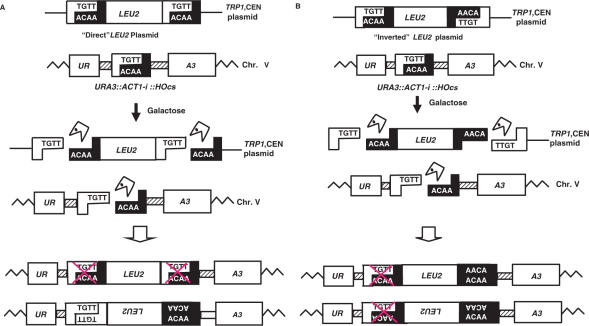
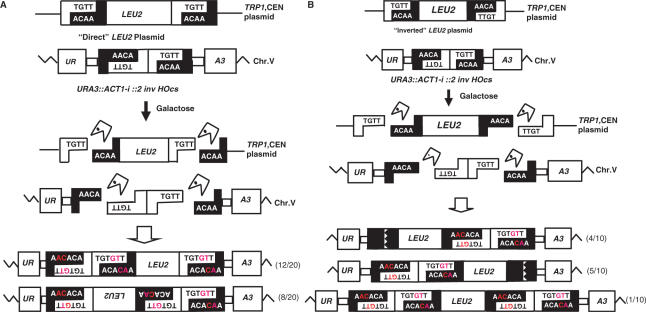
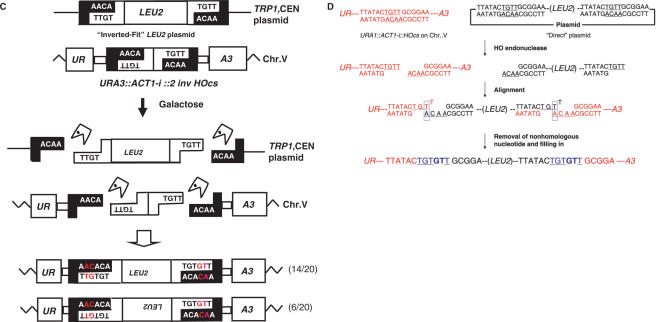
Double-strand breaks (DSBs) are dangerous chromosomal lesions that must be efficiently repaired in order to avoid loss of genetic information or cell death. In all organisms studied to date, two different mechanisms are used to repair DSBs: homologous recombination (HR) and non-homologous end joining (NHEJ). Previous studies have shown that during DSB repair, non-homologous exogenous DNA (also termed 'filler DNA') can be incorporated at the site of a DSB. We have created a genetic system in the yeast Saccharomyces cerevisiae to study the mechanism of fragment capture. Our yeast strains carry recognition sites for the HO endonuclease at a unique chromosomal site, and plasmids in which a LEU2 gene is flanked by HO cut sites. Upon induction of the HO endonuclease, a linear extrachromosomal fragment is generated in each cell and its incorporation at the chromosomal DSB site can be genetically monitored. Our results show that linear fragments are captured at the repaired DSB site at frequencies of 10(-6) to 10(-4) per plated cell depending on strain background and specific end sequences. The mechanism of fragment capture depends on the NHEJ machinery, but only partially on the homologous recombination proteins. More than one fragment can be used during repair, by a mechanism that relies on the annealing of small complementary sequences. We present a model to explain the basis for fragment capture.
Figures

Similar articles
-
Quantitation and analysis of the formation of HO-endonuclease stimulated chromosomal translocations by single-strand annealing in Saccharomyces cerevisiae.J Vis Exp. 2011 Sep 23;(55):3150. doi: 10.3791/3150. J Vis Exp. 2011. PMID: 21968396 Free PMC article.
-
Pathway utilization in response to a site-specific DNA double-strand break in fission yeast.EMBO J. 2003 Mar 17;22(6):1419-30. doi: 10.1093/emboj/cdg119. EMBO J. 2003. PMID: 12628934 Free PMC article.
-
The yeast chromatin remodeler RSC complex facilitates end joining repair of DNA double-strand breaks.Mol Cell Biol. 2005 May;25(10):3934-44. doi: 10.1128/MCB.25.10.3934-3944.2005. Mol Cell Biol. 2005. PMID: 15870268 Free PMC article.
-
Differential usage of non-homologous end-joining and homologous recombination in double strand break repair.DNA Repair (Amst). 2006 Sep 8;5(9-10):1021-9. doi: 10.1016/j.dnarep.2006.05.022. Epub 2006 Jun 27. DNA Repair (Amst). 2006. PMID: 16807135 Review.
-
A Life Investigating Pathways That Repair Broken Chromosomes.Annu Rev Genet. 2016 Nov 23;50:1-28. doi: 10.1146/annurev-genet-120215-035043. Epub 2016 Oct 3. Annu Rev Genet. 2016. PMID: 27732795 Review.
Cited by
-
RAD59 is required for efficient repair of simultaneous double-strand breaks resulting in translocations in Saccharomyces cerevisiae.DNA Repair (Amst). 2008 May 3;7(5):788-800. doi: 10.1016/j.dnarep.2008.02.003. Epub 2008 Mar 25. DNA Repair (Amst). 2008. PMID: 18373960 Free PMC article.
-
Mechanisms of change in gene copy number.Nat Rev Genet. 2009 Aug;10(8):551-64. doi: 10.1038/nrg2593. Nat Rev Genet. 2009. PMID: 19597530 Free PMC article. Review.
-
Diverse mechanisms of somatic structural variations in human cancer genomes.Cell. 2013 May 9;153(4):919-29. doi: 10.1016/j.cell.2013.04.010. Cell. 2013. PMID: 23663786 Free PMC article.
-
End-processing during non-homologous end-joining: a role for exonuclease 1.Nucleic Acids Res. 2011 Feb;39(3):970-8. doi: 10.1093/nar/gkq886. Epub 2010 Oct 8. Nucleic Acids Res. 2011. PMID: 20935051 Free PMC article.
-
Proofreading activity of DNA polymerase Pol2 mediates 3'-end processing during nonhomologous end joining in yeast.PLoS Genet. 2008 Apr 25;4(4):e1000060. doi: 10.1371/journal.pgen.1000060. PLoS Genet. 2008. PMID: 18437220 Free PMC article.
References
-
- Kupiec M. Damage-induced recombination in the yeast Saccharomyces cerevisiae. Mutat. Res. 2000;451:91–105. - PubMed
-
- Kupiec M, Steinlauf R. Damage-induced ectopic recombination in the yeast Saccharomyces cerevisiae. Mutat. Res. 1997;384:33–44. - PubMed
-
- Krogh BO, Symington LS. Recombination proteins in yeast. Annu. Rev. Genet. 2004;38:233–271. - PubMed
-
- Pastwa E, Blasiak J. Non-homologous DNA end joining. Acta Biochim. Pol. 2003;50:891–908. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Research Materials
