

Construction of a doxycycline-dependent simian immunodeficiency virus reveals a nontranscriptional function of tat in viral replication
- PMID: 17670816
- PMCID: PMC2045552
- DOI: 10.1128/JVI.01354-07
Construction of a doxycycline-dependent simian immunodeficiency virus reveals a nontranscriptional function of tat in viral replication
Abstract
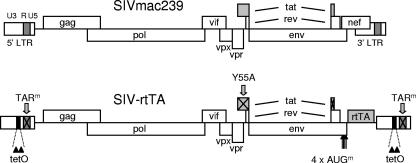
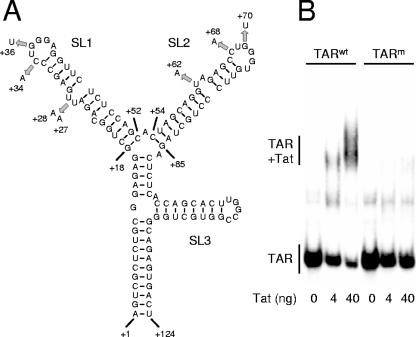
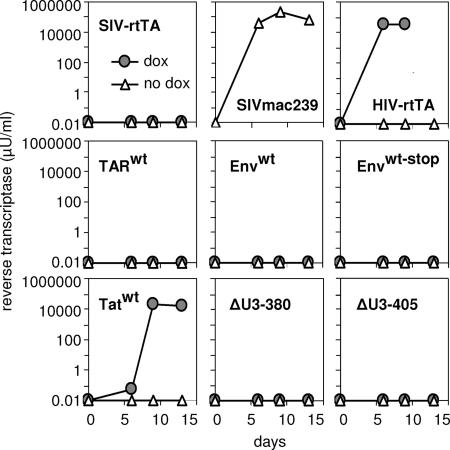
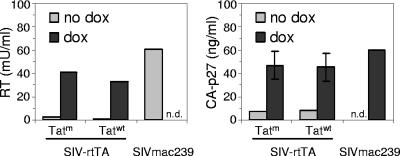
In the quest for an effective vaccine against human immunodeficiency virus (HIV), live attenuated virus vaccines have proven to be very effective in the experimental model system of simian immunodeficiency virus (SIV) in macaques. However, live attenuated HIV vaccines are considered unsafe for use in humans because the attenuated virus may accumulate genetic changes during persistence and evolve to a pathogenic variant. As an alternative approach, we earlier presented a conditionally live HIV-1 variant that replicates exclusively in the presence of doxycycline (DOX). Replication of this vaccine strain can be limited to the time that is needed to provide full protection through transient DOX administration. Since the effectiveness and safety of such a conditionally live AIDS vaccine should be tested in macaques, we constructed a similar DOX-dependent SIVmac239 variant in which the Tat-TAR (trans-acting responsive) transcription control mechanism was functionally replaced by the DOX-inducible Tet-On regulatory mechanism. Moreover, this virus can be used as a tool in SIV biology studies and vaccine research because both the level and duration of replication can be controlled by DOX administration. Unexpectedly, the new SIV variant required a wild-type Tat protein for replication, although gene expression was fully controlled by the incorporated Tet-On system. This result suggests that Tat has a second function in SIV replication in addition to its role in the activation of transcription.
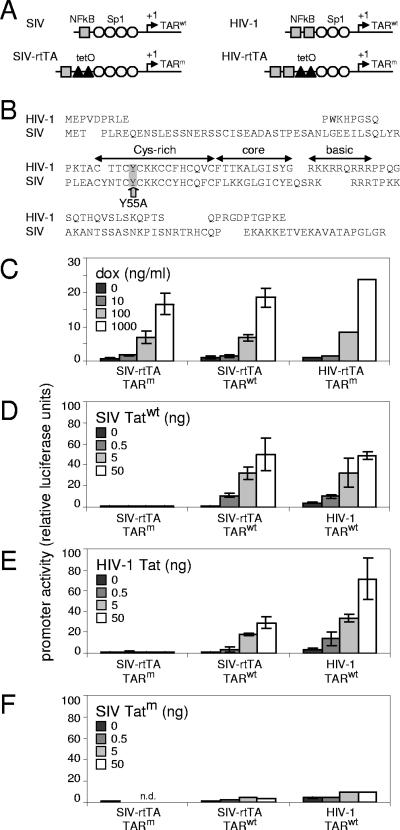
Figures
Similar articles
-
Optimization of the doxycycline-dependent simian immunodeficiency virus through in vitro evolution.Retrovirology. 2008 Jun 5;5:44. doi: 10.1186/1742-4690-5-44. Retrovirology. 2008. PMID: 18533993 Free PMC article.
-
Conditional virus replication as an approach to a safe live attenuated human immunodeficiency virus vaccine.J Neurovirol. 2002 Dec;8 Suppl 2:134-7. doi: 10.1080/13550280290101102. J Neurovirol. 2002. PMID: 12491165 Review.
-
Modification of the Tet-On regulatory system prevents the conditional-live HIV-1 variant from losing doxycycline-control.Retrovirology. 2006 Nov 9;3:82. doi: 10.1186/1742-4690-3-82. Retrovirology. 2006. PMID: 17094796 Free PMC article.
-
Dox-dependent SIVmac with tetracycline-inducible promoter in the U3 promoter region.Virology. 2000 Apr 10;269(2):268-75. doi: 10.1006/viro.2000.0213. Virology. 2000. PMID: 10753705
-
Conditionally replicating HIV and SIV variants.Virus Res. 2016 May 2;216:66-75. doi: 10.1016/j.virusres.2015.05.004. Epub 2015 May 15. Virus Res. 2016. PMID: 25982510 Review.
Cited by
-
Didehydro-Cortistatin A Inhibits HIV-1 by Specifically Binding to the Unstructured Basic Region of Tat.mBio. 2019 Feb 5;10(1):e02662-18. doi: 10.1128/mBio.02662-18. mBio. 2019. PMID: 30723126 Free PMC article.
-
The allosteric HIV-1 integrase inhibitor BI-D affects virion maturation but does not influence packaging of a functional RNA genome.PLoS One. 2014 Jul 29;9(7):e103552. doi: 10.1371/journal.pone.0103552. eCollection 2014. PLoS One. 2014. PMID: 25072705 Free PMC article.
-
Optimization of the doxycycline-dependent simian immunodeficiency virus through in vitro evolution.Retrovirology. 2008 Jun 5;5:44. doi: 10.1186/1742-4690-5-44. Retrovirology. 2008. PMID: 18533993 Free PMC article.
-
The Tat inhibitor didehydro-cortistatin A suppresses SIV replication and reactivation.FASEB J. 2019 Jul;33(7):8280-8293. doi: 10.1096/fj.201801165R. Epub 2019 Apr 25. FASEB J. 2019. PMID: 31021670 Free PMC article.
-
Tet-On Systems For Doxycycline-inducible Gene Expression.Curr Gene Ther. 2016;16(3):156-67. doi: 10.2174/1566523216666160524144041. Curr Gene Ther. 2016. PMID: 27216914 Free PMC article. Review.
References
-
- Auersperg, N. 1964. Long-term cultivation of hypodiploid human tumor cells. J. Natl. Cancer Inst. 32:135-163. - PubMed
-
- Baba, T. W., Y. S. Jeong, D. Penninck, R. Bronson, M. F. Greene, and R. M. Ruprecht. 1995. Pathogenicity of live, attenuated SIV after mucosal infection of neonatal macaques. Science 267:1820-1825. - PubMed
-
- Baba, T. W., V. Liska, A. H. Khimani, N. B. Ray, P. J. Dailey, D. Penninck, R. Bronson, M. F. Greene, H. M. McClure, L. N. Martin, and R. M. Ruprecht. 1999. Live attenuated, multiply deleted simian immunodeficiency virus causes AIDS in infant and adult macaques. Nat. Med. 5:194-203. - PubMed
-
- Baron, U., and H. Bujard. 2000. Tet repressor-based system for regulated gene expression in eukaryotic cells: principles and advances. Methods Enzymol. 327:401-421. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Research Materials
