

The interaction of APOBEC3G with human immunodeficiency virus type 1 nucleocapsid inhibits tRNA3Lys annealing to viral RNA
- PMID: 17670826
- PMCID: PMC2045548
- DOI: 10.1128/JVI.00162-07
The interaction of APOBEC3G with human immunodeficiency virus type 1 nucleocapsid inhibits tRNA3Lys annealing to viral RNA
Abstract
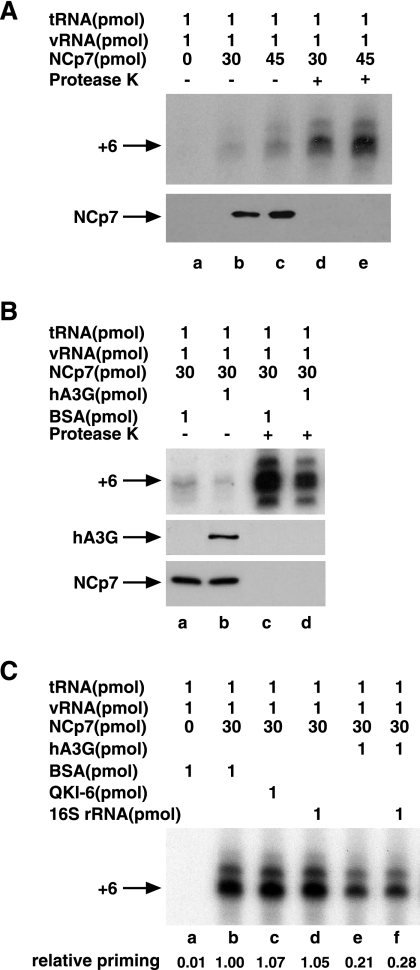
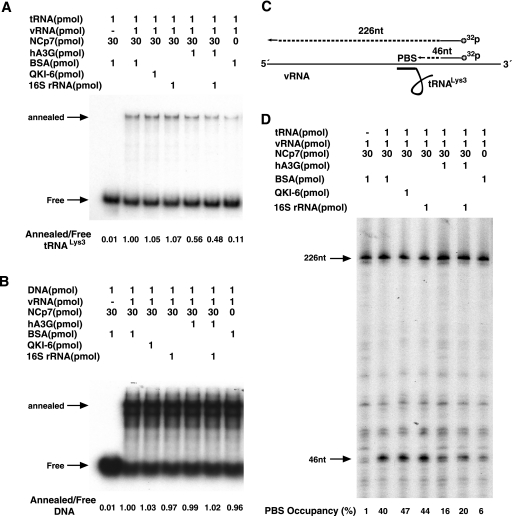
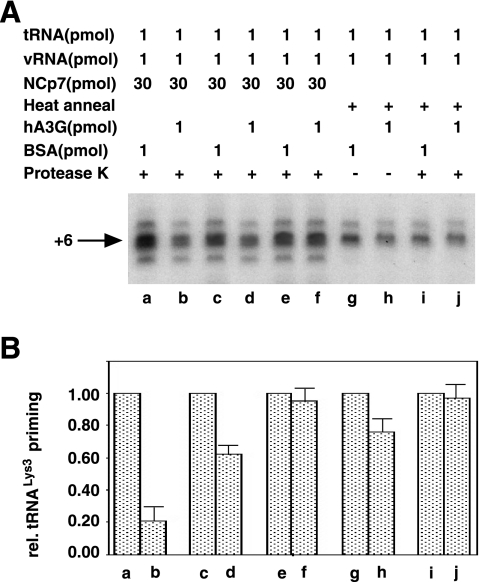
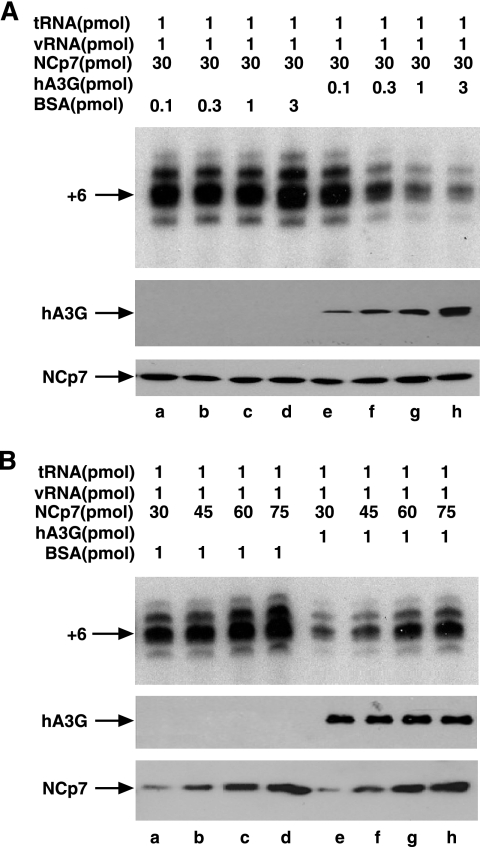
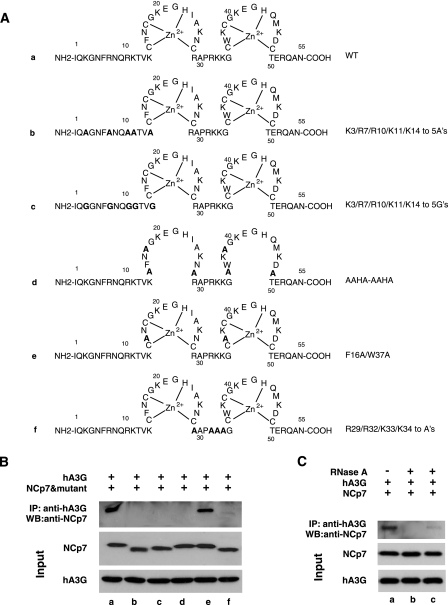
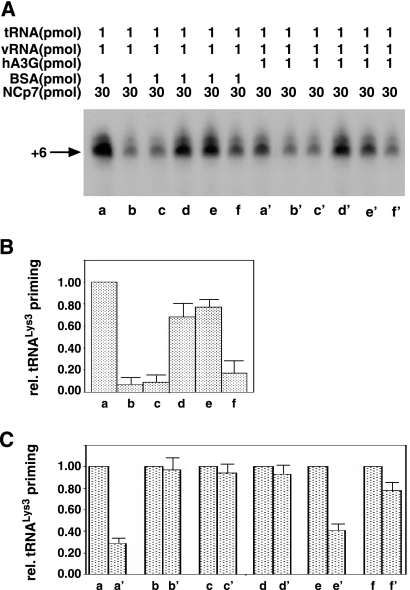
Human immunodeficiency virus type 1 (HIV-1) containing human APOBEC3G (hA3G) has a reduced ability to produce viral DNA in newly infected cells. At least part of this hA3G-facilitated inhibition is due to a cytidine deamination-independent reduction in the ability to initiate reverse transcription. HIV-1 nucleocapsid (NCp7) is required both for the incorporation of hA3G into virions and for the annealing between viral RNA and tRNA(3)(Lys), the primer tRNA for reverse transcription. Herein we present evidence that the interaction of hA3G with nucleocapsid is required for the inhibition of reverse transcription initiation. A tRNA(3)(Lys) priming complex was produced in vitro by the NCp7-facilitated annealing of tRNA(3)(Lys) to synthetic viral RNA in the absence or presence of hA3G. The effect of hA3G on the annealing of tRNA(3)(Lys) to viral RNA and the ability of tRNA(3)(Lys) to initiate reverse transcription was measured. Our results show the following. (i) Electrophoretic band shift and primer binding site assays show that hA3G reduces the annealing of tRNA(3)(Lys) 44 and 60%, respectively, but does not disrupt the annealed complex once formed. (ii) hA3G inhibits tRNA(3)(Lys) priming 70 to 80%. (iii) Inhibition of tRNA(3)(Lys) priming by hA3G requires an interaction between hA3G and NCp7 during annealing. Thus, annealing of tRNA(3)(Lys) is insensitive to hA3G inhibition when facilitated by a zinc finger mutant of NCp7 unable to interact with hA3G. NCp7-independent annealing of DNA to viral RNA also is insensitive to hA3G inhibition. These results indicate that hA3G does not sterically block tRNA(3)(Lys) annealing by binding to viral RNA. Annealing and priming are not affected by another RNA binding protein, QKI-6.
Figures
References
-
- Alce, T. M., and W. Popik. 2004. APOBEC3G is incorporated into virus-like particles by a direct interaction with HIV-1 Gag nucleocapsid protein. J. Biol. Chem. 279:34083-34086. - PubMed
-
- Arts, E. J., X. Li, Z. Gu, L. Kleiman, M. A. Parniak, and M. A. Wainberg. 1994. Comparison of deoxyoligonucleotide and tRNA(Lys-3) as primers in an endogenous human immunodeficiency virus-1 in vitro reverse transcription/template-switching reaction. J. Biol. Chem. 269:14672-14680. - PubMed
-
- Beerens, N., and B. Berkhout. 2000. In vitro studies on tRNA annealing and reverse transcription with mutant HIV-1 RNA templates. J. Biol. Chem. 275:15474-15481. - PubMed
-
- Briggs, J. A., M. N. Simon, I. Gross, H. G. Krausslich, S. D. Fuller, V. M. Vogt, and M. C. Johnson. 2004. The stoichiometry of Gag protein in HIV-1. Nat. Struct. Mol. Biol. 11:672-675. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
