

Alterations in receptor binding properties of recent human influenza H3N2 viruses are associated with reduced natural killer cell lysis of infected cells
- PMID: 17670834
- PMCID: PMC2045558
- DOI: 10.1128/JVI.01217-07
Alterations in receptor binding properties of recent human influenza H3N2 viruses are associated with reduced natural killer cell lysis of infected cells
Abstract
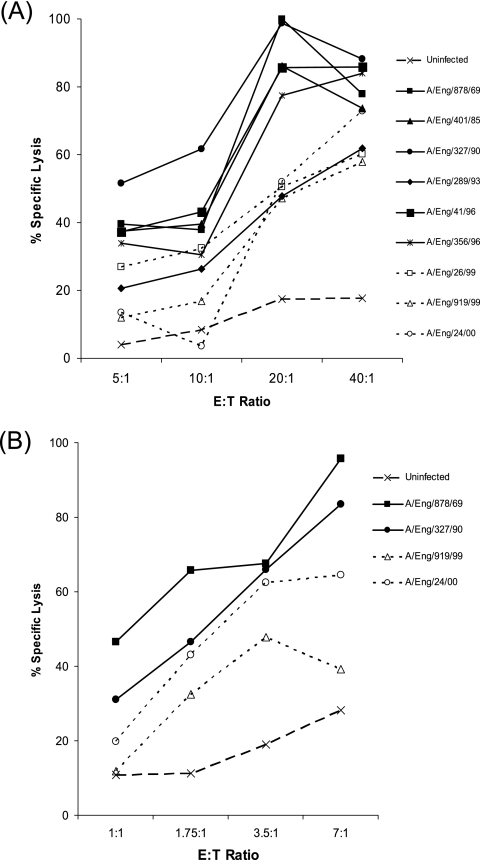
Natural killer (NK) cell recognition of influenza virus-infected cells involves hemagglutinin (HA) binding to sialic acid (SA) on activating NK receptors. SA also acts as a receptor for the binding of influenza virus to its target host cells. The SA binding properties of H3N2 influenza viruses have been observed to change during circulation in humans: recent isolates are unable to agglutinate chicken red blood cells and show reduced affinity for synthetic glycopolymers representing SA-alpha-2,3-lactose (3'SL-PAA) and SA-alpha-2,6-N-acetyl lactosamine (6'SLN-PAA) carbohydrates. Here, NK lysis of cells infected with human H3N2 influenza viruses isolated between 1969 and 2003 was analyzed. Cells infected with recent isolates (1999 to 2003) were found to be lysed less effectively than cells infected with older isolates (1969 to 1996). This change occurred concurrently with the acquisition of two new potential glycosylation site motifs in HA. Deletion of the potential glycosylation site motif at 133 to 135 in HA1 from a recent isolate partially restored the agglutination phenotype to a recombinant virus, indicating that the HA-SA interaction is inhibited by the glycosylation modification. Deletion of either of the recently acquired potential glycosylation sites from HA led to increased NK lysis of cells infected with recombinant viruses carrying modified HA. These results indicate that alterations in HA glycosylation may affect NK cell recognition of influenza virus-infected cells in addition to virus binding to host cells.
Figures
Similar articles
-
Structural and evolutionary characteristics of HA, NA, NS and M genes of clinical influenza A/H3N2 viruses passaged in human and canine cells.J Clin Virol. 2009 Aug;45(4):322-33. doi: 10.1016/j.jcv.2009.05.030. J Clin Virol. 2009. PMID: 19546028
-
Probing of the receptor-binding sites of the H1 and H3 influenza A and influenza B virus hemagglutinins by synthetic and natural sialosides.Virology. 1993 Sep;196(1):111-21. doi: 10.1006/viro.1993.1459. Virology. 1993. PMID: 8356788
-
Avian influenza A viruses differ from human viruses by recognition of sialyloligosaccharides and gangliosides and by a higher conservation of the HA receptor-binding site.Virology. 1997 Jun 23;233(1):224-34. doi: 10.1006/viro.1997.8580. Virology. 1997. PMID: 9201232
-
Tumor and viral recognition by natural killer cells receptors.Semin Cancer Biol. 2006 Oct;16(5):348-58. doi: 10.1016/j.semcancer.2006.07.005. Epub 2006 Jul 7. Semin Cancer Biol. 2006. PMID: 16893656 Review.
-
[Receptor sialylsugar chains as determinants of host range of influenza viruses].Nihon Rinsho. 2000 Nov;58(11):2206-10. Nihon Rinsho. 2000. PMID: 11225305 Review. Japanese.
Cited by
-
HA Antibody-Mediated FcγRIIIa Activity Is Both Dependent on FcR Engagement and Interactions between HA and Sialic Acids.Front Immunol. 2016 Sep 29;7:399. doi: 10.3389/fimmu.2016.00399. eCollection 2016. Front Immunol. 2016. PMID: 27746785 Free PMC article.
-
Receptor specificity and erythrocyte binding preferences of avian influenza viruses isolated from India.Virol J. 2012 Oct 30;9:251. doi: 10.1186/1743-422X-9-251. Virol J. 2012. PMID: 23110802 Free PMC article.
-
Early responses of natural killer cells in pigs experimentally infected with 2009 pandemic H1N1 influenza A virus.PLoS One. 2014 Jun 23;9(6):e100619. doi: 10.1371/journal.pone.0100619. eCollection 2014. PLoS One. 2014. PMID: 24955764 Free PMC article.
-
High efficiency gene transfer to airways of mice using influenza hemagglutinin pseudotyped lentiviral vectors.J Gene Med. 2013 Jan;15(1):51-62. doi: 10.1002/jgm.2695. J Gene Med. 2013. PMID: 23319179 Free PMC article.
-
Differential activation of NK cells by influenza A pseudotype H5N1 and 1918 and 2009 pandemic H1N1 viruses.J Virol. 2010 Aug;84(15):7822-31. doi: 10.1128/JVI.00069-10. Epub 2010 May 19. J Virol. 2010. PMID: 20484512 Free PMC article.
References
-
- Abed, Y., A. M. Bourgault, R. J. Fenton, P. J. Morley, D. Gower, I. J. Owens, M. Tisdale, and G. Boivin. 2002. Characterisation of two influenza A (H3N2) clinical isolates with reduced susceptibility to neuraminidase inhibitors due to mutations in the haemagglutinin gene. J. Infect. Dis. 186:1074-1080. - PubMed
-
- Achdout, H., T. I. Arnon, G. Markel, T. Gonen-Gross, G. Katz, N. Lieberman, R. Gazit, A. Joseph, E. Kedar, and O. Mandelboim. 2003. Enhanced recognition of human NK receptors after influenza virus infection. J. Immunol. 171:915-923. - PubMed
-
- Arnon, T. I., H. Achdout, N. Lieberman, R. Gazit, T. Gonen-Gross, G. Katz, A. Bar-Ilan, N. Bloushtain, M. Lev, A. Joseph, E. Kedar, A. Porgador, and O. Mandelboim. 2004. The mechanisms controlling the recognition of tumor- and virus-infected cells by NKp46. Blood 103:664-672. - PubMed
-
- Arnon, T. I., M. Lev, G. Katz, Y. Chernobrov, A. Porgador, and O. Mandelboim. 2001. Recognition of viral hemagglutinins by NKp44 but not by NKp30. Eur. J. Immunol. 31:2680-2689. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Medical
Research Materials
