

NMDA di-heteromeric receptor populations and associated proteins in rat hippocampus
- PMID: 17670980
- PMCID: PMC2263005
- DOI: 10.1523/JNEUROSCI.2155-07.2007
NMDA di-heteromeric receptor populations and associated proteins in rat hippocampus
Abstract
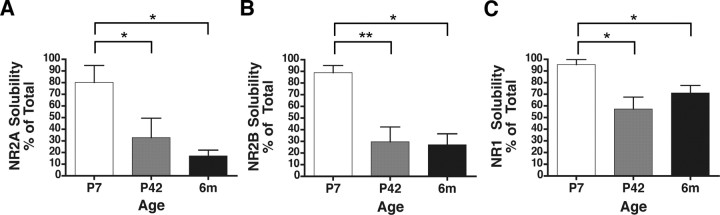
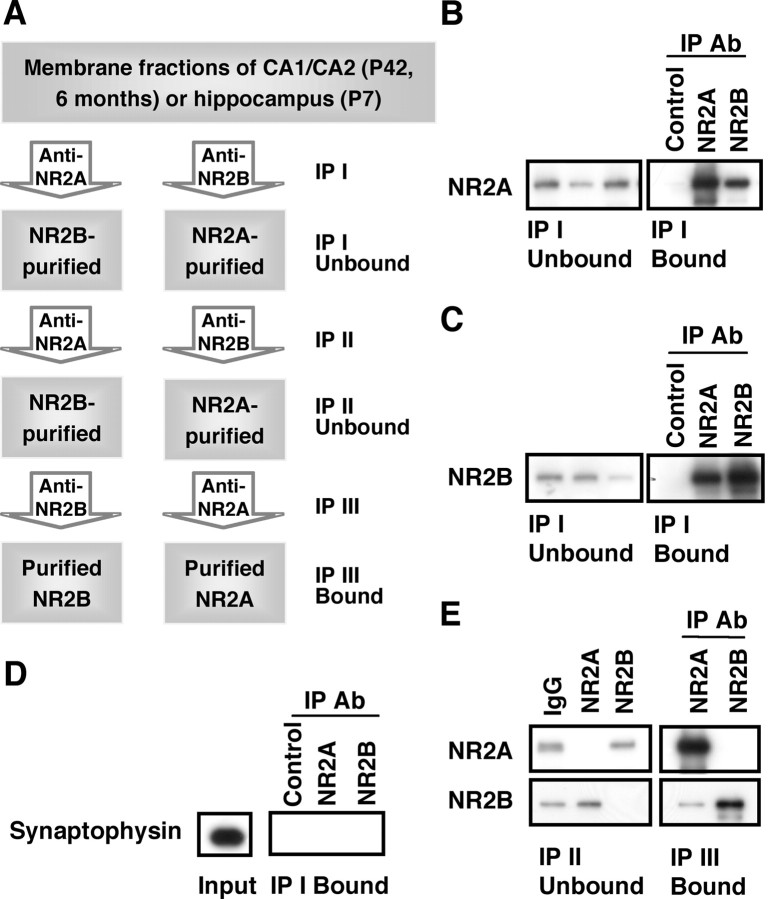
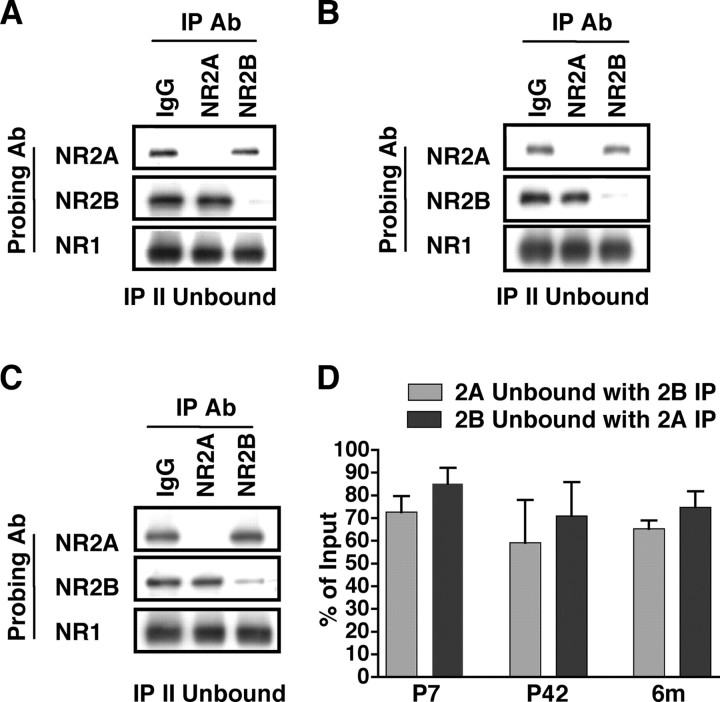
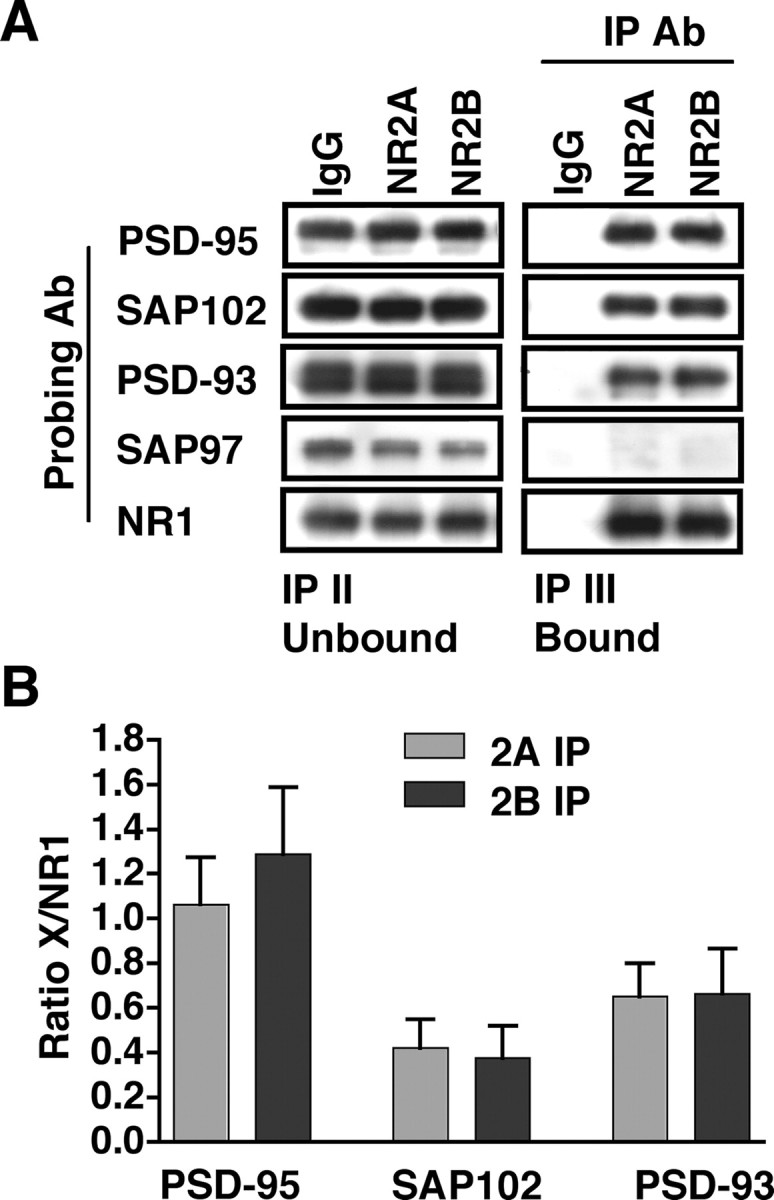
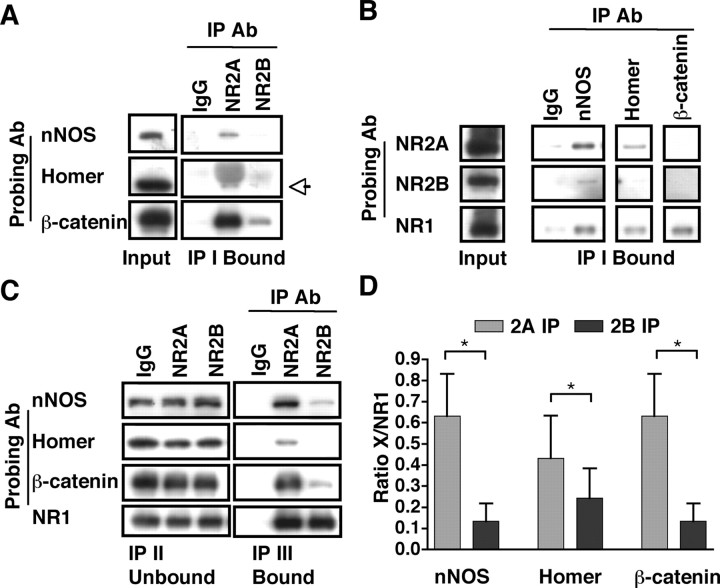
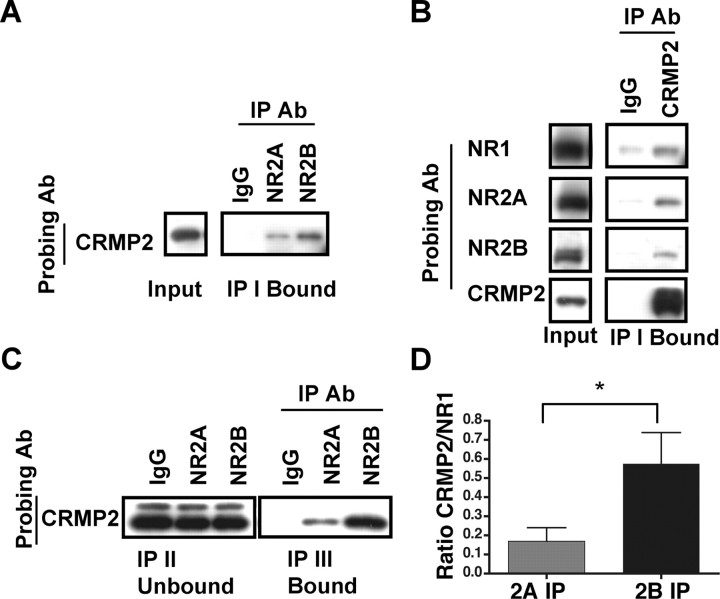
Subunit composition of NMDA receptors (NMDARs) determines a range of physiological properties, downstream signaling effects, and binding partners. Differential localization of NR2A- or NR2B-containing NMDARs within the neuron and subunit-specific protein associations may explain differences in NR2A and NR2B contributions to synaptic plasticity and excitotoxic cell death. This question is complicated by the existence of tri-heteromeric complexes (NR1/NR2A/NR2B). To date, no quantitative biochemical determinations have been made of the relative abundance of different NMDAR populations in intact hippocampus, the region extensively correlated with NMDAR-dependent long-term potentiation. We investigated subunit composition and subunit-specific interactions in CA1/CA2 of rat hippocampus. Using sequential immunoprecipitations to deplete either NR2B or NR2A, di-heteromeric NR1/NR2A and NR1/NR2B receptor populations were isolated from postnatal day 7 (P7) hippocampus and P42 and 6-month-old CA1/CA2. Quantitative Western blot analysis revealed that 60-70% of NR2A and 70-85% of NR2B subunits were associated in NR1/NR2A or NR1/NR2B di-heteromeric complexes. Isolated di-heteromeric receptor fractions were used to examine NR2A- or NR2B-specific interactions with synapse-associated proteins. Our results indicate that NR2A- or NR2B-containing NMDARs associate similarly with postsynaptic density-95 (PSD-95), synapse-associated protein 102, and PSD-93 at P42. However, NR2A-containing receptors coimmunoprecipitated a greater proportion of the synaptic proteins neuronal nitric oxide synthase, Homer, and beta-catenin. Finally, mass spectrometry analysis of isolated di-heteromeric receptors identified a novel NMDAR interactor, collapsin response mediator protein 2, which preferentially associates with NR2B-containing di-heteromeric NMDARs. In summary, in rat hippocampus, NR2A and NR2B exist primarily in di-heteromeric complexes that interact similarly with PSD-95-related proteins but are associated with different protein complexes.
Figures
References
-
- Al-Hallaq RA, Jarabek BR, Fu Z, Vicini S, Wolfe BB, Yasuda RP. Association of NR3A with the N-methyl-d-aspartate receptor NR1 and NR2 subunits. Mol Pharmacol. 2002;62:1119–1127. - PubMed
-
- Besshoh S, Bawa D, Teves L, Wallace MC, Gurd JW. Increased phosphorylation and redistribution of NMDA receptors between synaptic lipid rafts and post-synaptic densities following transient global ischemia in the rat brain. J Neurochem. 2005;93:186–194. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Miscellaneous