

Olfactory bulb gamma oscillations are enhanced with task demands
- PMID: 17670982
- PMCID: PMC6673062
- DOI: 10.1523/JNEUROSCI.1199-07.2007
Olfactory bulb gamma oscillations are enhanced with task demands
Abstract
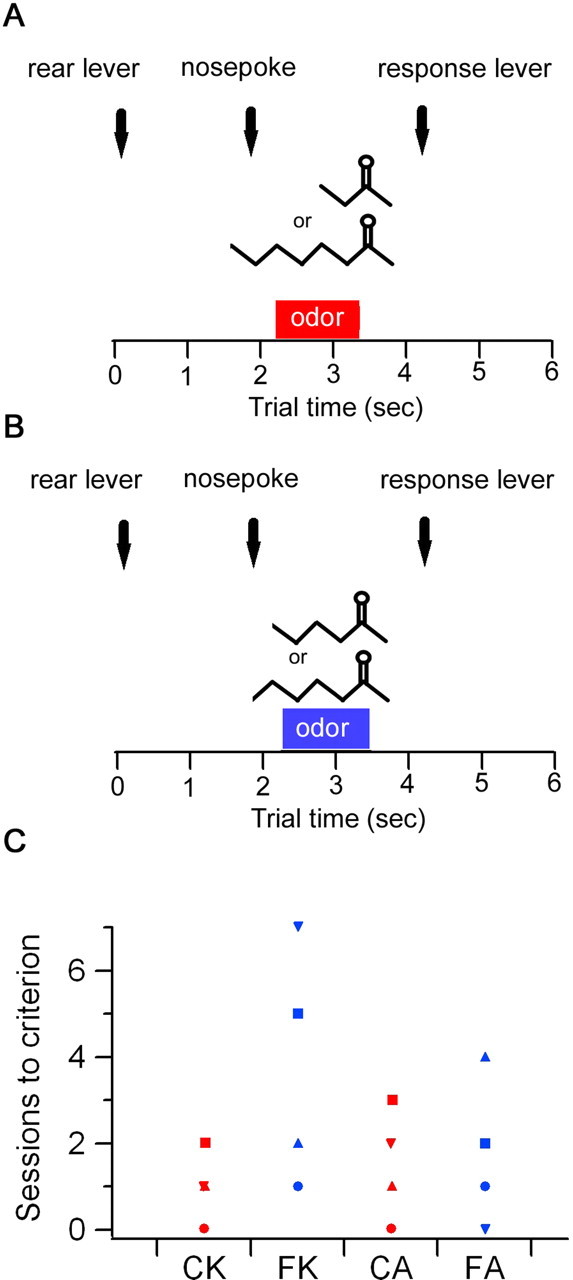
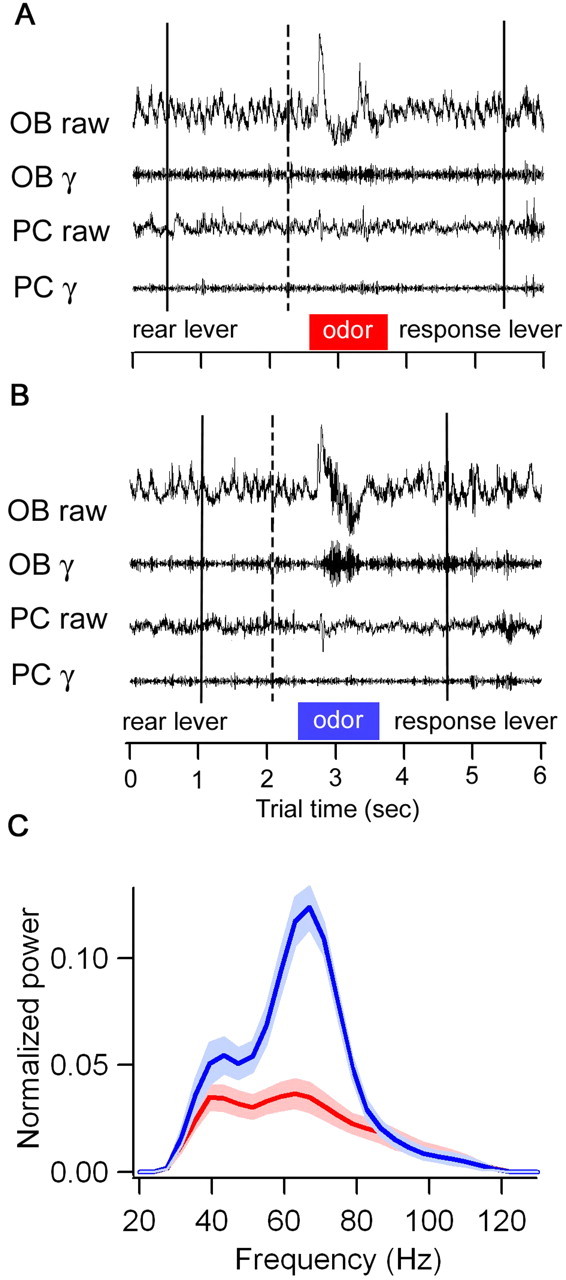
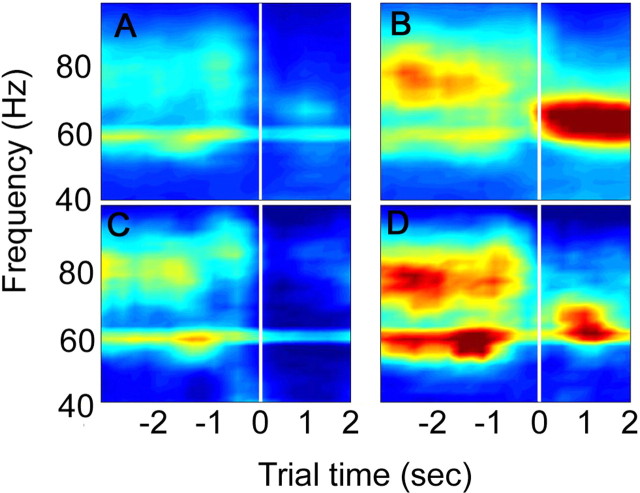
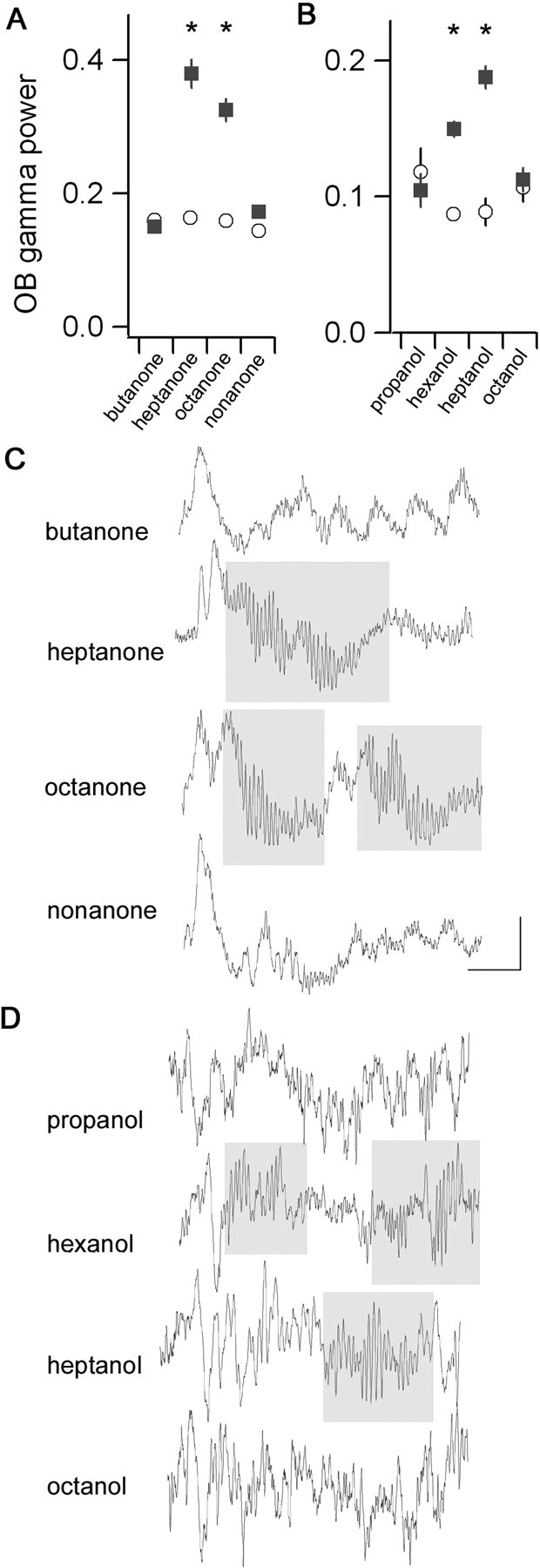
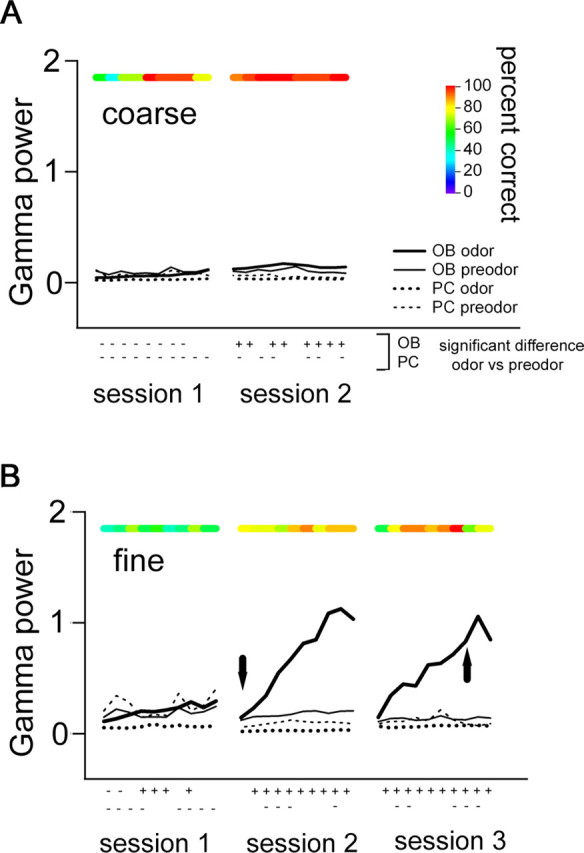
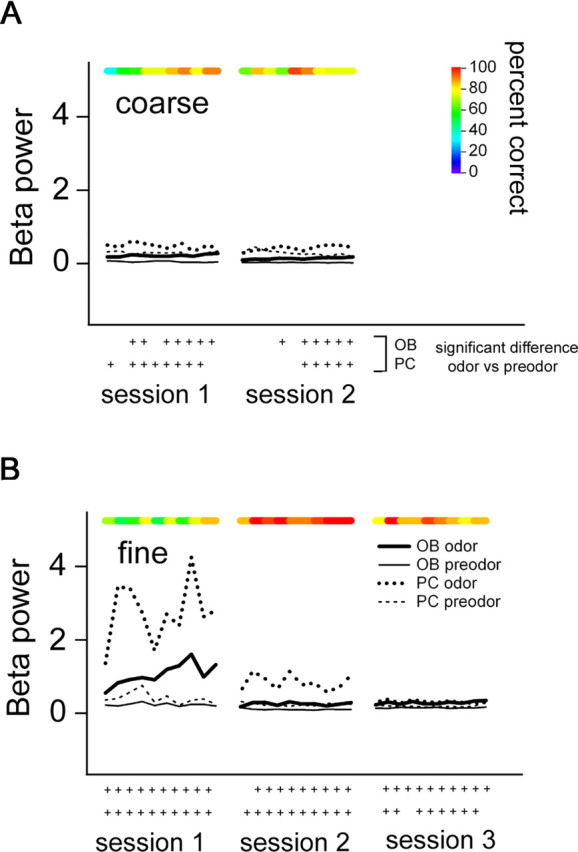
Fast oscillations in neural assemblies have been proposed as a mechanism to facilitate stimulus representation in a variety of sensory systems across animal species. In the olfactory system, intervention studies suggest that oscillations in the gamma frequency range play a role in fine odor discrimination. However, there is still no direct evidence that such oscillations are intrinsically altered in intact systems to aid in stimulus disambiguation. Here we show that gamma oscillatory power in the rat olfactory bulb during a two-alternative choice task is modulated in the intact system according to task demands with dramatic increases in gamma power during discrimination of molecularly similar odorants in contrast to dissimilar odorants. This elevation in power evolves over the course of criterion performance, is specific to the gamma frequency band (65-85 Hz), and is independent of changes in the theta or beta frequency band range. Furthermore, these high amplitude gamma oscillations are restricted to the olfactory bulb, such that concurrent piriform cortex recordings show no evidence of enhanced gamma power during these high-amplitude events. Our results display no modulation in the power of beta oscillations (15-28 Hz) shown previously to increase with odor learning in a Go/No-go task, and we suggest that the oscillatory profile of the olfactory system may be influenced by both odor discrimination demands and task type. The results reported here indicate that enhancement of local gamma power may reflect a switch in the dynamics of the system to a strategy that optimizes stimulus resolution when input signals are ambiguous.
Figures
References
-
- Abraham NM, Spors H, Carleton A, Margrie TW, Kuner T, Schaefer AT. Maintaining accuracy at the expense of speed: stimulus similarity defines odor discrimination time in mice. Neuron. 2004;44:865–876. - PubMed
-
- Bressler SL, Freeman WJ. Frequency analysis of olfactory system EEG in cat, rabbit, and rat. Electroencephalogr Clin Neurophysiol. 1980;50:19–24. - PubMed
Publication types
MeSH terms
Grants and funding
LinkOut - more resources
Full Text Sources