

J-protein co-chaperone Sis1 required for generation of [RNQ+] seeds necessary for prion propagation
- PMID: 17673909
- PMCID: PMC1952226
- DOI: 10.1038/sj.emboj.7601811
J-protein co-chaperone Sis1 required for generation of [RNQ+] seeds necessary for prion propagation
Abstract
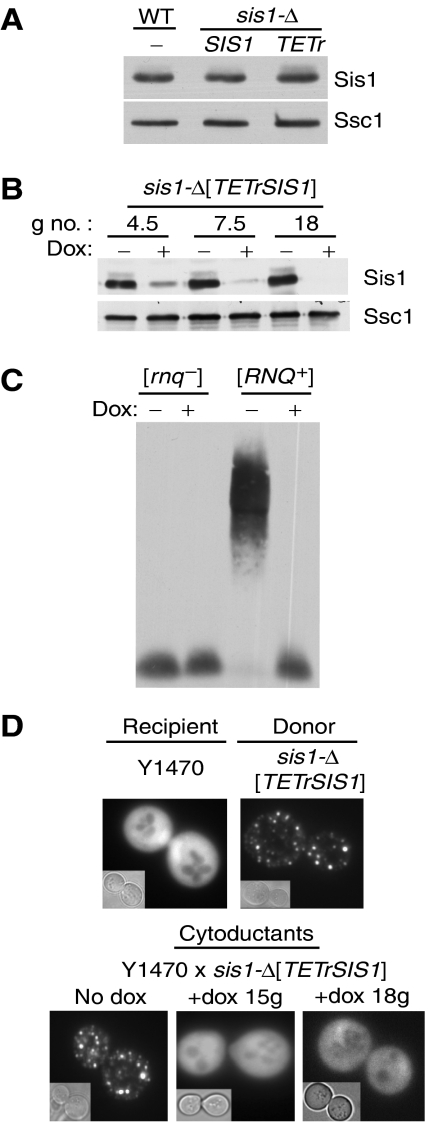
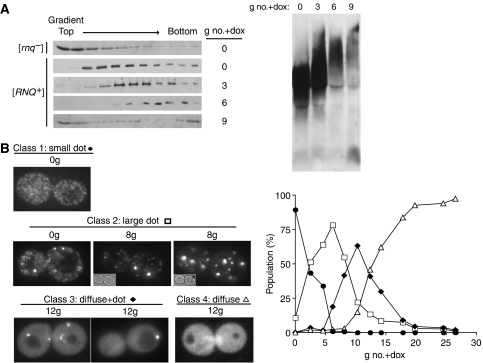
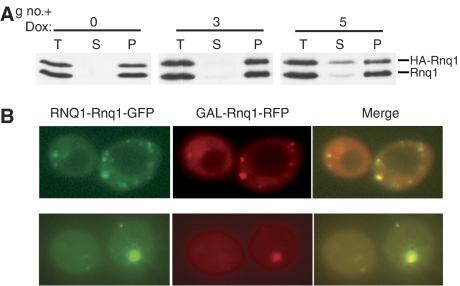
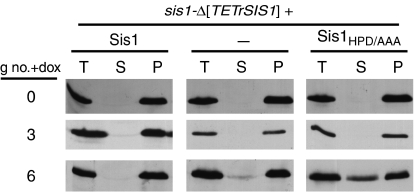
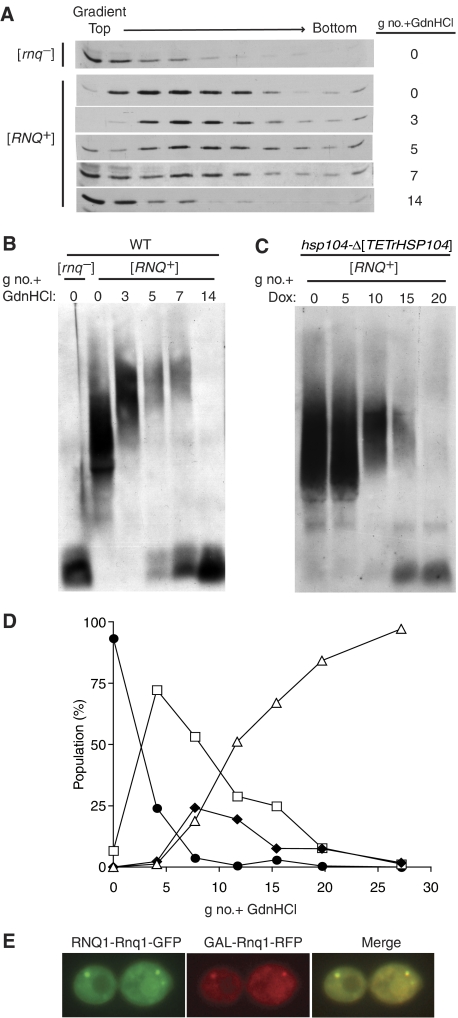
Yeast prions are protein-based genetic elements capable of self-perpetuation. One such prion, [RNQ(+)], requires the J-protein Sis1, an Ssa Hsp70 co-chaperone, as well as the AAA+ ATPase, Hsp104, for its propagation. We report that, upon depletion of Sis1, as well as upon inactivation of Hsp104, Rnq1 aggregates increased in size. Subsequently, cells having large aggregates, as well as an apparently soluble pool of Rnq1, became predominant in the cell population. Newly synthesized Rnq1 localized to both aggregates and bulk cytosol, suggesting that nascent Rnq1 partitioned into pools of prion and nonprion conformations, and implying that these large aggregates were still active as seeds. Ultimately, soluble Rnq1 predominated, and the prion was lost from the population. Our data suggest a model in which J-protein:Hsp70 machinery functions in prion propagation, in conjunction with Hsp104. Together, these chaperones facilitate fragmentation of prion polymers, generating a sufficient number of seeds to allow efficient conversion of newly synthesized Rnq1 into the prion conformation.
Figures
References
-
- Bagriantsev S, Liebman SW (2004) Specificity of prion assembly in vivo. [PSI+] and [PIN+] form separate structures in yeast. J Biol Chem 279: 51042–51048 - PubMed
-
- Borchsenius AS, Muller S, Newnam GP, Inge-Vechtomov SG, Chernoff YO (2006) Prion variant maintained only at high levels of the Hsp104 disaggregase. Curr Genet 49: 21–29 - PubMed
-
- Bosl B, Grimminger V, Walter S (2006) The molecular chaperone Hsp104—a molecular machine for protein disaggregation. J Struct Biol 156: 139–148 - PubMed
-
- Bradley ME, Bagriantsev S, Vishveshwara N, Liebman SW (2003) Guanidine reduces stop codon read-through caused by missense mutations in SUP35 or SUP45. Yeast 20: 625–632 - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
Research Materials
