

Kinesin is an evolutionarily fine-tuned molecular ratchet-and-pawl device of decisively locked direction
- PMID: 17675343
- PMCID: PMC2072077
- DOI: 10.1529/biophysj.107.108233
Kinesin is an evolutionarily fine-tuned molecular ratchet-and-pawl device of decisively locked direction
Abstract
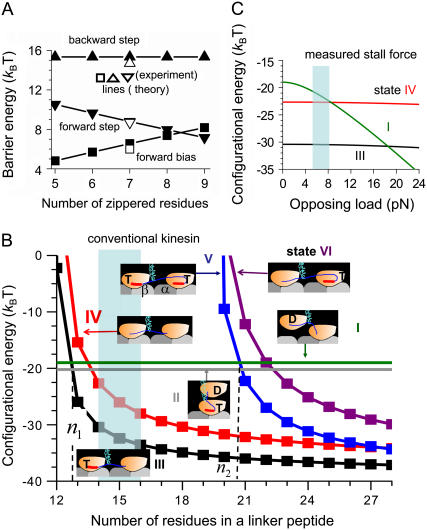
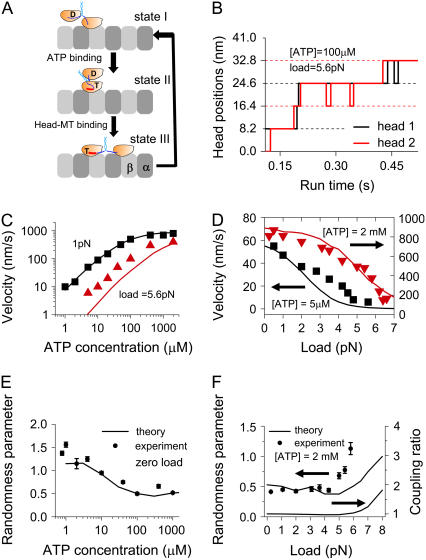
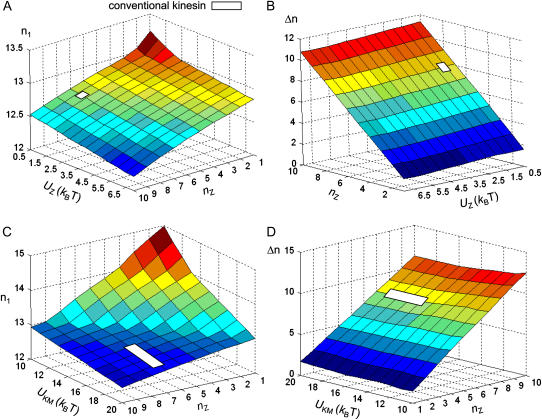
Conventional kinesin is a dimeric motor protein that transports membranous organelles toward the plus-end of microtubules (MTs). Individual kinesin dimers show steadfast directionality and hundreds of consecutive steps, yet the detailed physical mechanism remains unclear. Here we compute free energies for the entire dimer-MT system for all possible interacting configurations by taking full account of molecular details. Employing merely first principles and several measured binding and barrier energies, the system-level analysis reveals insurmountable energy gaps between configurations, asymmetric ground state caused by mechanically lifted configurational degeneracy, and forbidden transitions ensuring coordination between both motor domains for alternating catalysis. This wealth of physical effects converts a kinesin dimer into a molecular ratchet-and-pawl device, which determinedly locks the dimer's movement into the MT plus-end and ensures consecutive steps in hand-over-hand gait. Under a certain range of extreme loads, however, the ratchet-and-pawl device becomes defective but not entirely abolished to allow consecutive back-steps. This study yielded quantitative evidence that kinesin's multiple molecular properties have been evolutionarily adapted to fine-tune the ratchet-and-pawl device so as to ensure the motor's distinguished performance.
Figures
Similar articles
-
The intrinsic load-resisting capacity of kinesin.Phys Biol. 2009 Apr 15;6(3):036002. doi: 10.1088/1478-3975/6/3/036002. Phys Biol. 2009. PMID: 19369705
-
Modeling motility of the kinesin dimer from molecular properties of individual monomers.Biochemistry. 2008 Apr 22;47(16):4733-42. doi: 10.1021/bi800072p. Epub 2008 Mar 28. Biochemistry. 2008. PMID: 18370409
-
The E-hook of tubulin interacts with kinesin's head to increase processivity and speed.Biophys J. 2005 Nov;89(5):3223-34. doi: 10.1529/biophysj.104.057505. Epub 2005 Aug 12. Biophys J. 2005. PMID: 16100283 Free PMC article.
-
Kinesin's moonwalk.Curr Opin Cell Biol. 2006 Feb;18(1):61-7. doi: 10.1016/j.ceb.2005.12.009. Epub 2005 Dec 19. Curr Opin Cell Biol. 2006. PMID: 16361092 Review.
-
Kinesin: a molecular motor with a spring in its step.Proc Biol Sci. 2002 Nov 22;269(1507):2363-71. doi: 10.1098/rspb.2002.2117. Proc Biol Sci. 2002. PMID: 12495505 Free PMC article. Review.
Cited by
-
Autonomous DNA molecular motor tailor-designed to navigate DNA origami surface for fast complex motion and advanced nanorobotics.Sci Adv. 2023 Sep 22;9(38):eadi8444. doi: 10.1126/sciadv.adi8444. Epub 2023 Sep 22. Sci Adv. 2023. PMID: 37738343 Free PMC article.
-
Mechanism of processive movement of monomeric and dimeric kinesin molecules.Int J Biol Sci. 2010 Nov 3;6(7):665-74. doi: 10.7150/ijbs.6.665. Int J Biol Sci. 2010. PMID: 21060728 Free PMC article. Review.
-
Polymer-Based Accurate Positioning: An Exact Worm-like-Chain Study.ACS Omega. 2018 Oct 29;3(10):14318-14326. doi: 10.1021/acsomega.8b01448. eCollection 2018 Oct 31. ACS Omega. 2018. PMID: 31458122 Free PMC article.
-
Chemically Fueled Self-Assembly in Biology and Chemistry.Angew Chem Int Ed Engl. 2021 Sep 6;60(37):20120-20143. doi: 10.1002/anie.202100274. Epub 2021 Apr 7. Angew Chem Int Ed Engl. 2021. PMID: 33704885 Free PMC article. Review.
-
How Well Can DNA Rupture DNA? Shearing and Unzipping Forces inside DNA Nanostructures.ACS Omega. 2018 Jan 31;3(1):292-301. doi: 10.1021/acsomega.7b01692. Epub 2018 Jan 10. ACS Omega. 2018. PMID: 30023776 Free PMC article.
References
-
- Vale, R. D., and R. A. Milligan. 2000. The way things move: looking under the hood of molecular motor proteins. Science. 288:88–95. - PubMed
-
- Visscher, K., M. J. Schnitzer, and S. M. Block. 1999. Single kinesin molecules studied with a molecular force clamp. Nature. 400:184–189. - PubMed
-
- Carter, N. J., and R. A. Cross. 2005. Mechanics of the kinesin step. Nature. 435:308–312. - PubMed
-
- Schnitzer, M. J., K. Visscher, and S. M. Block. 2000. Force production by single kinesin motors. Nat. Cell Biol. 2:718–723. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
