

A mycobacterium ESX-1-secreted virulence factor with unique requirements for export
- PMID: 17676952
- PMCID: PMC1937011
- DOI: 10.1371/journal.ppat.0030105
A mycobacterium ESX-1-secreted virulence factor with unique requirements for export
Abstract
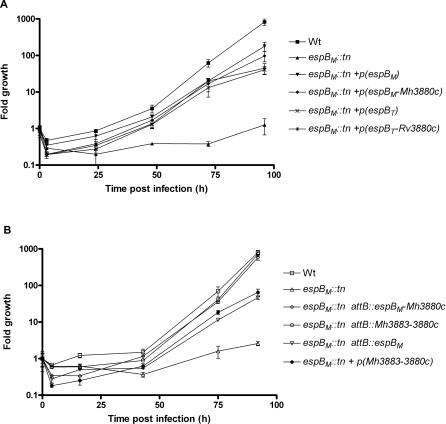
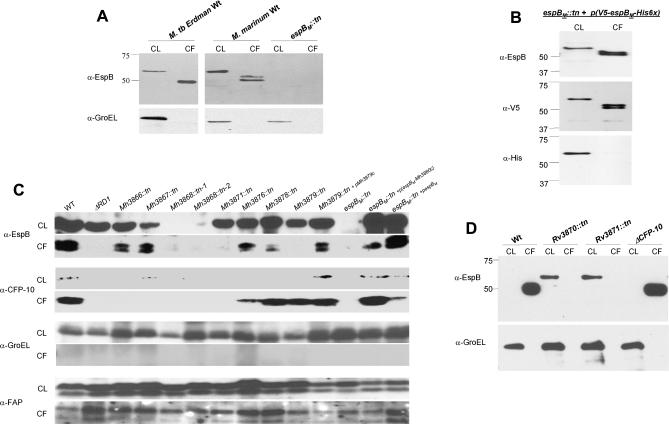
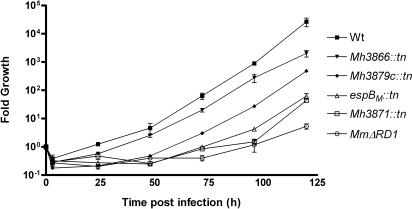
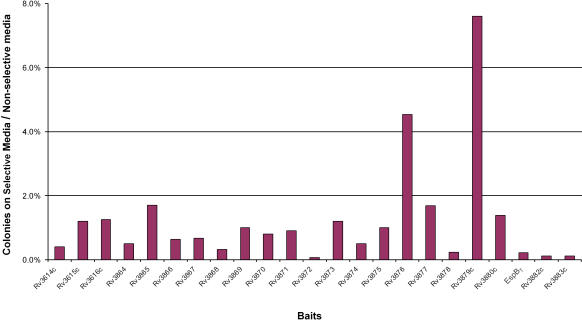
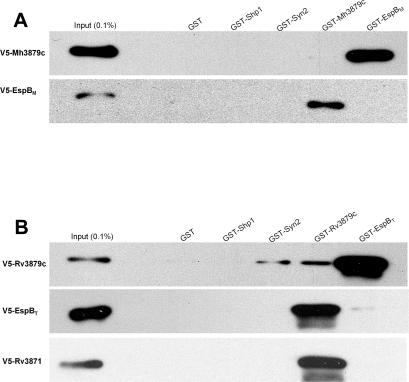
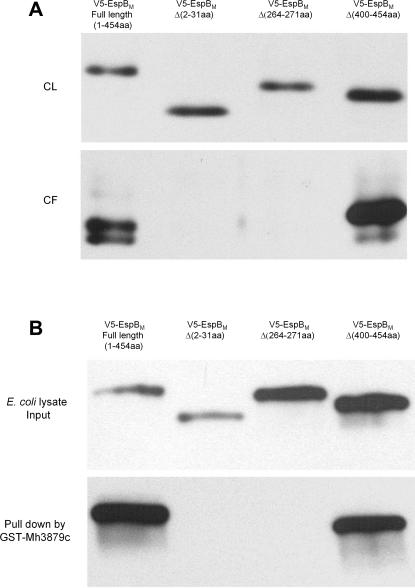
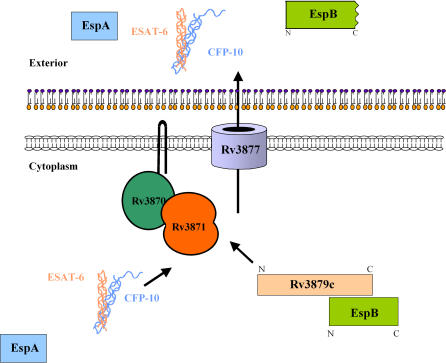
Specialized secretion systems of pathogenic bacteria commonly transport multiple effectors that act in concert to control and exploit the host cell as a replication-permissive niche. Both the Mycobacterium marinum and the Mycobacterium tuberculosis genomes contain an extended region of difference 1 (extRD1) locus that encodes one such pathway, the early secretory antigenic target 6 (ESAT-6) system 1 (ESX-1) secretion apparatus. ESX-1 is required for virulence and for secretion of the proteins ESAT-6, culture filtrate protein 10 (CFP-10), and EspA. Here, we show that both Rv3881c and its M. marinum homolog, Mh3881c, are secreted proteins, and disruption of RD1 in either organism blocks secretion. We have renamed the Rv3881c/Mh3881c gene espB for ESX-1 substrate protein B. Secretion of M. marinum EspB (EspBM) requires both the Mh3879c and Mh3871 genes within RD1, while CFP-10 secretion is not affected by disruption of Mh3879c. In contrast, disruption of Mh3866 or Mh3867 within the extRD1 locus prevents CFP-10 secretion without effect on EspBM. Mutants that fail to secrete only EspBM or only CFP-10 are less attenuated in macrophages than mutants failing to secrete both substrates. EspBM physically interacts with Mh3879c; the M. tuberculosis homolog, EspBT, physically interacts with Rv3879c; and mutants of EspBM that fail to bind Mh3879c fail to be secreted. We also found interaction between Rv3879c and Rv3871, a component of the ESX-1 machine, suggesting a mechanism for the secretion of EspB. The results establish EspB as a substrate of ESX-1 that is required for virulence and growth in macrophages and suggests that the contribution of ESX-1 to virulence may arise from the secretion of multiple independent substrates.
Conflict of interest statement
Figures
References
-
- Brodin P, Rosenkrands I, Andersen P, Cole ST, Brosch R. ESAT-6 proteins: Protective antigens and virulence factors? Trends Microbiol. 2004;12:500–508. - PubMed
Publication types
MeSH terms
Substances
Associated data
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Miscellaneous
