

Review
doi: 10.1146/annurev.psych.59.103006.093544.
The biological basis of audition
Affiliations
- PMID: 17678445
- PMCID: PMC3856181
- DOI: 10.1146/annurev.psych.59.103006.093544
Item in Clipboard
Review
The biological basis of audition
Annu Rev Psychol.
2008.
Abstract
Interest has recently surged in the neural mechanisms of audition, particularly with regard to functional imaging studies in human subjects. This review emphasizes recent work on two aspects of auditory processing. The first explores auditory spatial processing and the role of the auditory cortex in both nonhuman primates and human subjects. The interactions with visual stimuli, the ventriloquism effect, and the ventriloquism aftereffect are also reviewed. The second aspect is temporal processing. Studies investigating temporal integration, forward masking, and gap detection are reviewed, as well as examples from the birdsong system and echolocating bats.
Figures
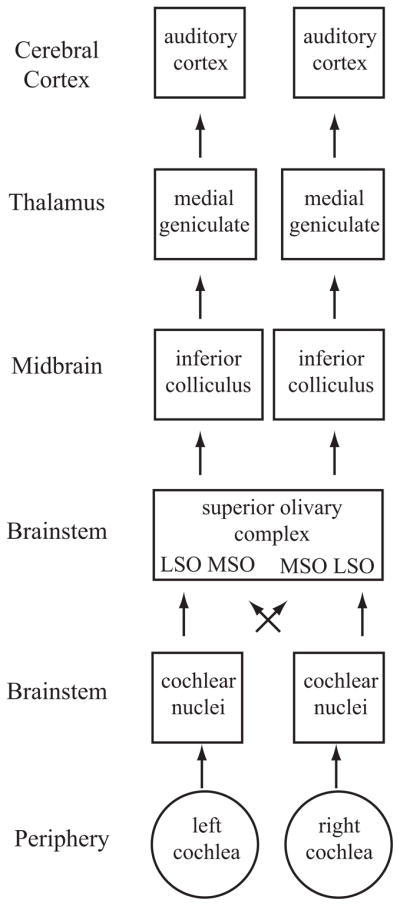
Schematic diagram of the ascending auditory system. Shown are the major subcortical nuclei that process acoustic information. Each cochlea projects to the ipsilateral cochlear nucleus in the brainstem. Not shown are the three different subdivisions of this nucleus, the dorsal cochlear nucleus, the anterior ventral cochlear nucleus, and the posterior ventral cochlear nucleus. These neurons project to the superior olivary complex (SOC) in the brainstem, which contains the lateral superior olive (LSO) and medial superior olive (MSO). Each cochlear nucleus projects to the SOC bilaterally; thus, neurons in these regions have binaural response properties that are projected upward to each ascending nucleus. The nucleus of the trapezoid body is not shown. SOC neurons project to the inferior colliculus (IC), which itself is composed of several subnuclei. The IC is the obligatory relay nucleus to the thalamus. Not shown is the nucleus of the lateral lemniscus. The IC neurons project to the medial geniculate body of the dorsal thalamus, which in turn projects to the auditory cortex.
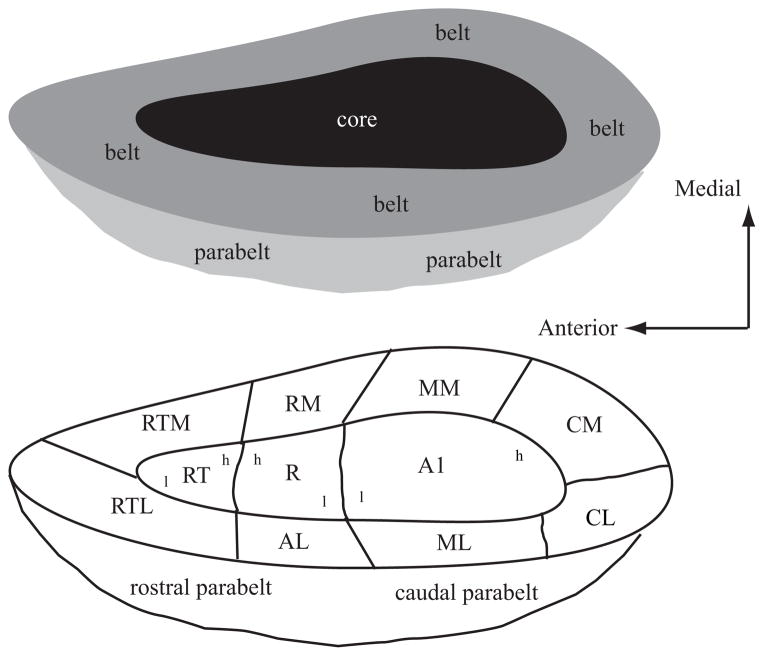
Schematic diagram of the primate auditory cortex. Top schematic shows the general organization of core (black), belt ( gray), and parabelt (light gray). Bottom schematic shows the approximate locations of the different cortical areas. The core is made up of the primary auditory cortex (AI), the rostral field (R), and the rostrotemporal (RT) area. These three fields have a tonotopic organization as shown by the lower-case letters h (high frequency) and l (low frequency). The medial belt includes the caudomedial (CM) area, middle medial (MM) area, rostromedial (RM) area, and medial rostrotemporal (RTM) area. The lateral belt includes the caudolateral (CL) area, middle lateral (ML) area, anterolateral (AL) area, and the lateral rostrotemporal (RTL) area. The parabelt is divided into rostral and caudal areas. Drawing is not to scale as the medial belt is significantly narrower than the lateral belt. Adapted from (Kaas & Hackett 2000).
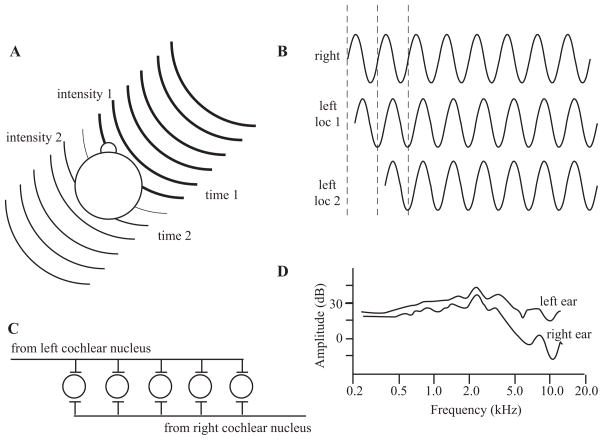
Sound localization cues. (A) Interaural intensity and timing cues. The head and body will absorb some of the sound energy, resulting in a lower-intensity stimulus at the far ear (intensity 2) in comparison with the near ear (intensity 1), represented here by the lines becoming thinner. The wave fronts will also arrive at the near ear (time 1) before the far ear (time 2), giving rise to interaural timing cues. (B) Interaural time and phase cues. The waveform will reach the far ear at a later time than the near ear. Ambiguities exist when only considering the phase and not the onset, as two locations can have the same phase disparity if they differ by multiples of a single cycle. (C ) Jeffress’s model of computing interaural timing differences. The lengths of the axons will result in coincident activation of only one neuron, depending on the location of the stimulus. (D) Head-related transfer functions. The acoustic energy measured at the eardrum varies between the two ears as a function of the stimulus location. (D) is adapted from (Wightman & Kistler 1989a).
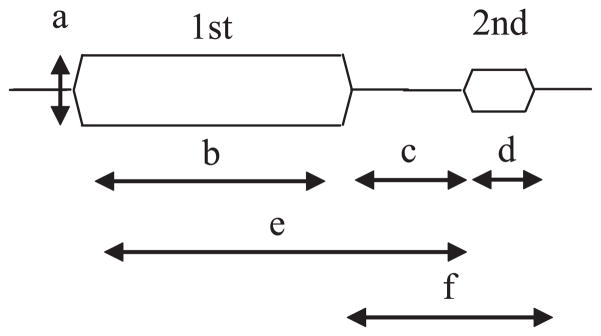
Stimulus parameters in forward masking experiments. The first stimulus is the masker and the subject is asked to detect the second stimulus. Parameters that influence the strength of masking include the intensity of the masker (a), the duration of the masker (b), the interstimulus interval (c), the duration of the target (d ), the onset interval between the masker and target (e), and the combined interstimulus interval and target duration ( f ).
References
-
- Ahissar M, Ahissar E, Bergman H, Vaadia E. Encoding of sound-source location and movement: activity of single neurons and interactions between adjacent neurons in the monkey auditory cortex. J Neurophysiol. 1992;67:203–15. - PubMed
-
- Alais D, Burr D. The ventriloquist effect results from near-optimal bimodal integration. Curr Biol. 2004;14:257–62. - PubMed
-
- Altmann CF, Bledowski C, Wibral M, Kaiser J. Processing of location and pattern changes of natural sounds in the human auditory cortex. NeuroImage. 2007;35:1192–200. - PubMed
-
- Altshuler MW, Comalli PE. Effect of stimulus intensity and frequency on median horizontal plane sound localization. J Aud Res. 1975;15:262–65.
Publication types
MeSH terms
Grants and funding
LinkOut - more resources
Full Text Sources
Miscellaneous
