

Wnt/Axin1/beta-catenin signaling regulates asymmetric nodal activation, elaboration, and concordance of CNS asymmetries
- PMID: 17678853
- PMCID: PMC1940036
- DOI: 10.1016/j.neuron.2007.07.007
Wnt/Axin1/beta-catenin signaling regulates asymmetric nodal activation, elaboration, and concordance of CNS asymmetries
Abstract
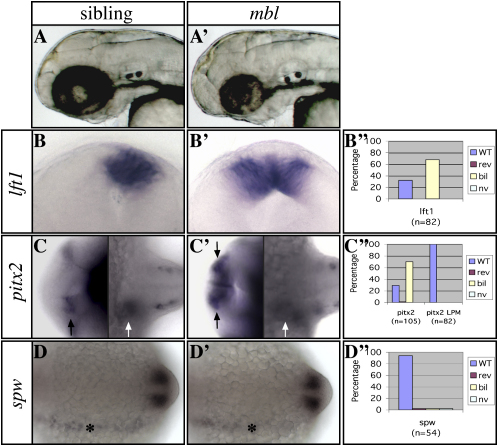
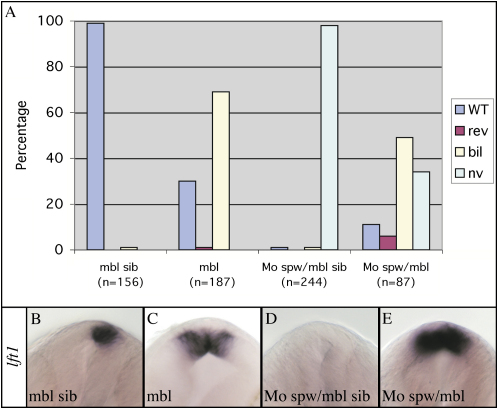
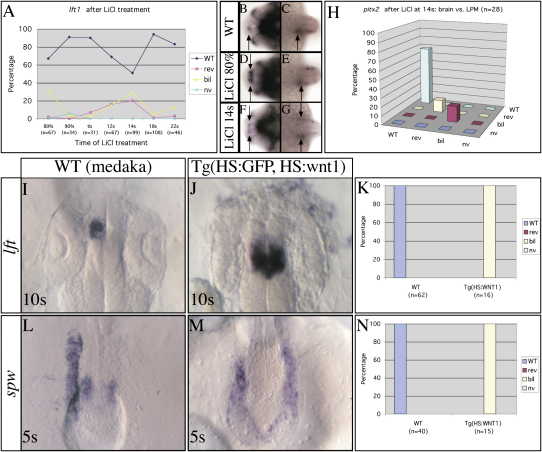
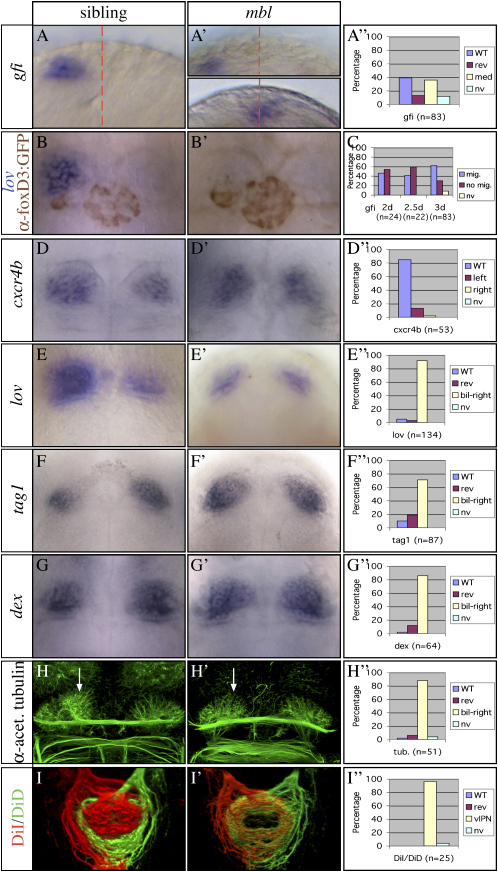
Nodal activity in the left lateral plate mesoderm (LPM) is required to activate left-sided Nodal signaling in the epithalamic region of the zebrafish forebrain. Epithalamic Nodal signaling subsequently determines the laterality of neuroanatomical asymmetries. We show that overactivation of Wnt/Axin1/beta-catenin signaling during late gastrulation leads to bilateral epithalamic expression of Nodal pathway genes independently of LPM Nodal signaling. This is consistent with a model whereby epithalamic Nodal signaling is normally bilaterally repressed, with Nodal signaling from the LPM unilaterally alleviating repression. We suggest that Wnt signaling regulates the establishment of the bilateral repression. We identify a second role for the Wnt pathway in the left/right regulation of LPM Nodal pathway gene expression, and finally, we show that at later stages Axin1 is required for the elaboration of concordant neuroanatomical asymmetries.
Figures
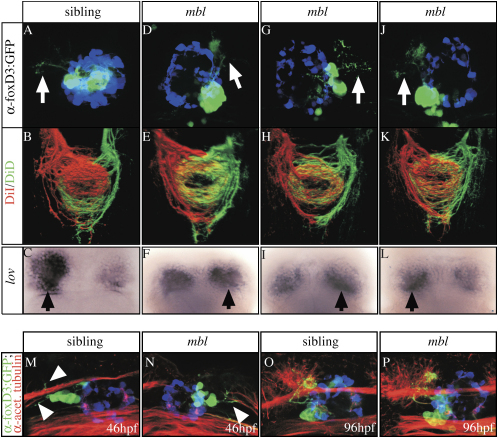
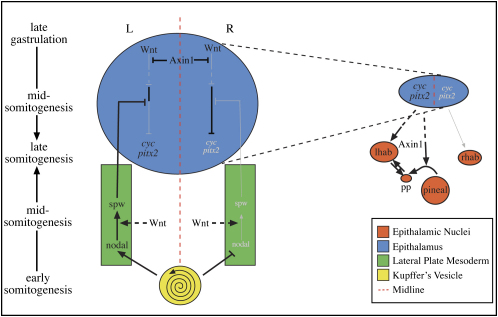
References
-
- Aghaallaei N., Bajoghli B., Walter I., Czerny T. Duplicated members of the Groucho/Tle gene family in fish. Dev. Dyn. 2005;234:143–150. - PubMed
-
- Aizawa H., Goto M., Sato T., Okamoto H. Temporally regulated asymmetric neurogenesis causes left-right difference in the zebrafish habenular structures. Dev. Cell. 2007;12:87–98. - PubMed
-
- Amack J.D., Yost H.J. The T box transcription factor no tail in ciliated cells controls zebrafish left-right asymmetry. Curr. Biol. 2004;14:685–690. - PubMed
-
- Bajoghli B., Aghaallaei N., Soroldoni D., Czerny T. The roles of Groucho/Tle in left-right asymmetry and Kupffer's vesicle organogenesis. Dev. Biol. 2007;303:347–361. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
- BB/C51454X/1/BB_/Biotechnology and Biological Sciences Research Council/United Kingdom
- G19181/BB_/Biotechnology and Biological Sciences Research Council/United Kingdom
- G0501487/MRC_/Medical Research Council/United Kingdom
- G0501487(76596)/MRC_/Medical Research Council/United Kingdom
- 074376/WT_/Wellcome Trust/United Kingdom
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
