

Tissue inhibitor of metalloproteinase-2 (TIMP-2) regulates myogenesis and beta1 integrin expression in vitro
- PMID: 17678891
- PMCID: PMC2197161
- DOI: 10.1016/j.yexcr.2007.06.007
Tissue inhibitor of metalloproteinase-2 (TIMP-2) regulates myogenesis and beta1 integrin expression in vitro
Abstract
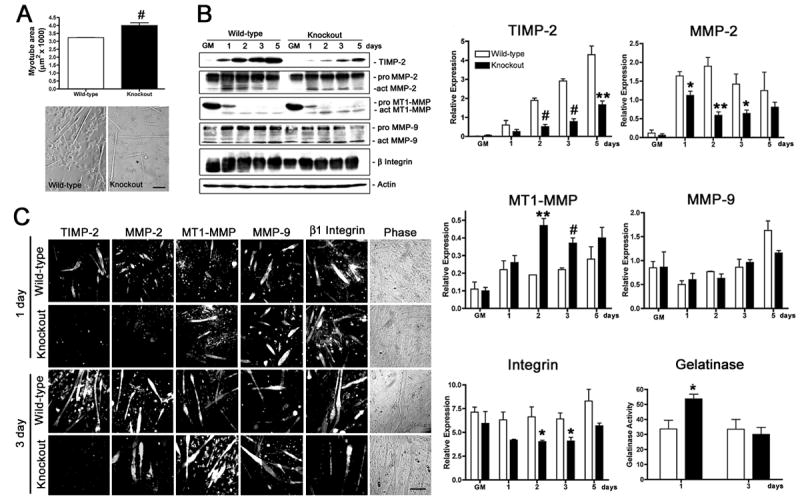
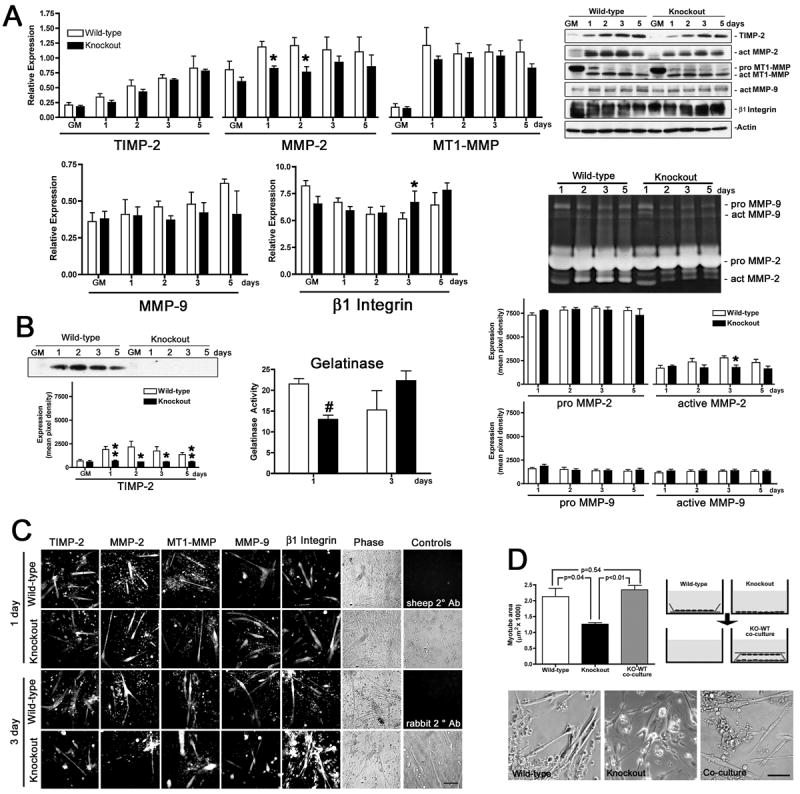
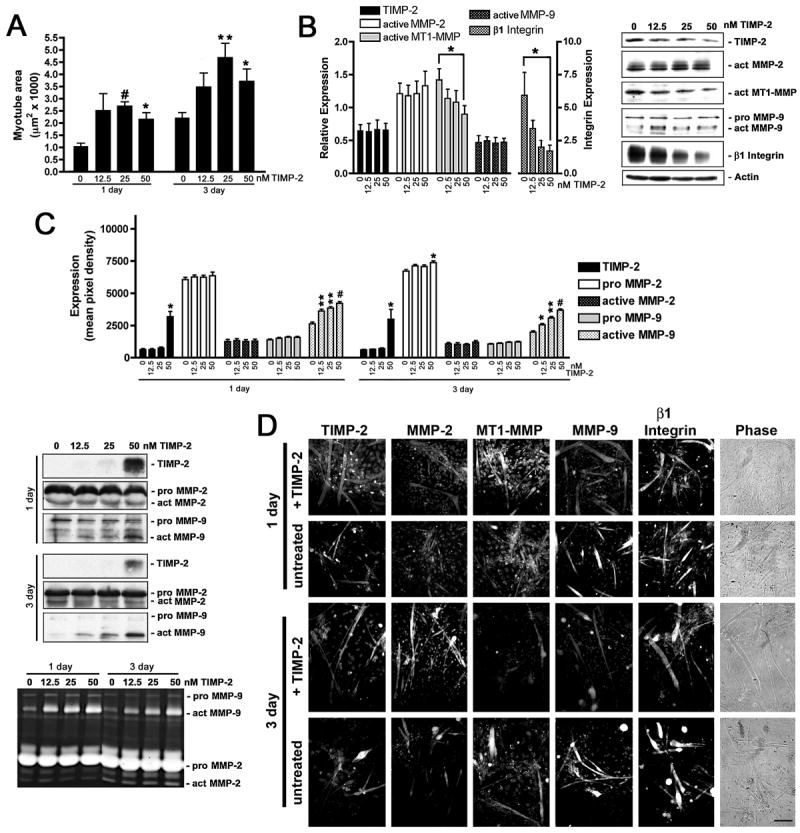
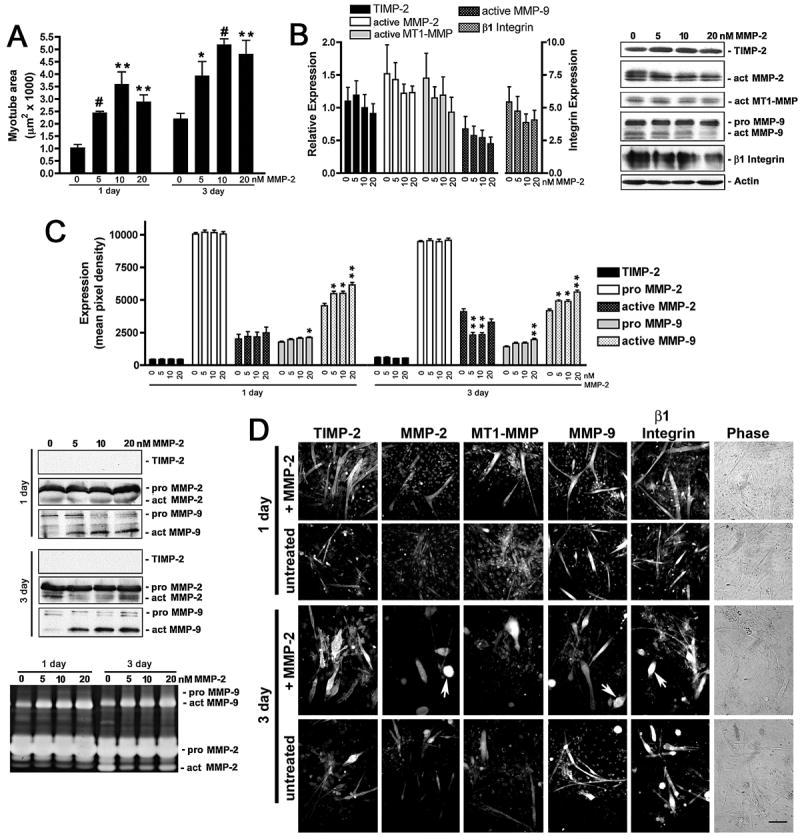
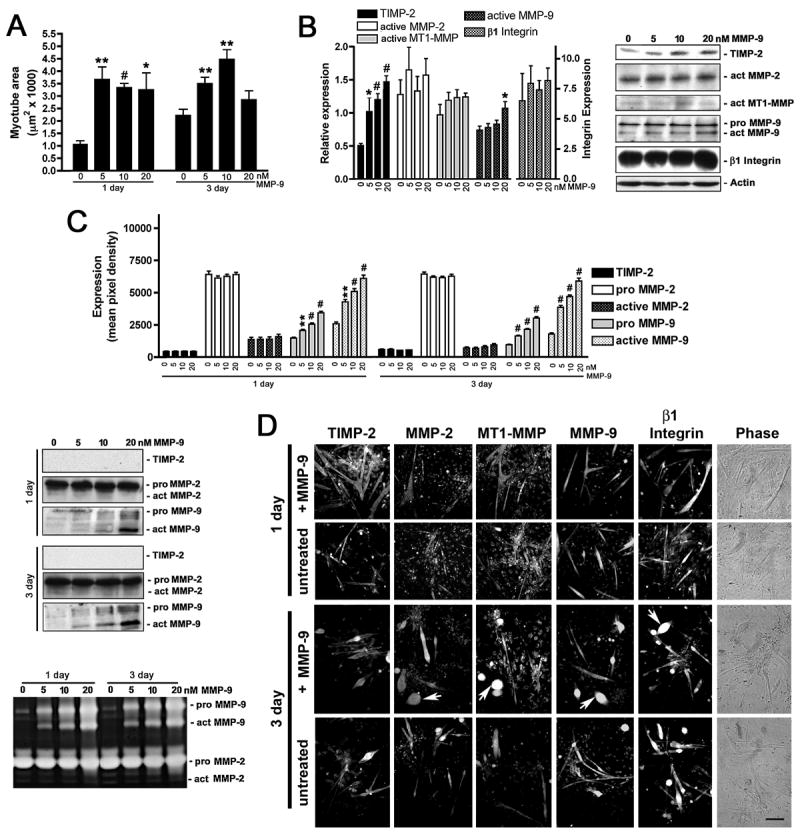
Myogenesis in vitro involves myoblast cell cycle arrest, migration, and fusion to form multinucleated myotubes. Extracellular matrix (ECM) integrity during these processes is maintained by the opposing actions of matrix metalloproteinase (MMP) proteases and their inhibitors, the tissue inhibitor of metalloproteinases (TIMPs). Here, we report that TIMP-2, MMP-2, and MT1-MMP are differentially expressed during mouse myoblast differentiation in vitro. A specific role for TIMP-2 in myogenesis is demonstrated by altered TIMP-2(-/-) myotube formation. When differentiated in horse serum-containing medium, TIMP-2(-/-) myotubes are larger than wild-type myotubes. In contrast, when serum-free medium is used, TIMP-2(-/-) myotubes are smaller than wild-type myotubes. Regardless of culture condition, myotube size is directly correlated with MMP activity and inversely correlated with beta1 integrin expression. Treatment with recombinant TIMP-2 rescues reduced TIMP-2(-/-) myotube size and induces increased MMP-9 activation and decreased beta1 integrin expression. Treatment with either MMP-2 or MMP-9 similarly rescues reduced myotube size, but has no effect on beta1 integrin expression. These data suggest a specific regulatory relationship between TIMP-2 and beta1 integrin during myogenesis. Elucidating the role of TIMP-2 in myogenesis in vitro may lead to new therapeutic options for the use of TIMP-2 in myopathies and muscular dystrophies in vivo.
Figures
References
-
- Parker MH, Seale P, Rudnicki MA. Looking back to the embryo: defining transcriptional networks in adult myogenesis. Nat Rev Genet. 2003;4:497–507. - PubMed
-
- Brand-Saberi B. Genetic and epigenetic control of skeletal muscle development. Ann Anat. 2005;187:199–207. - PubMed
-
- Krauss RS, Cole F, Gaio U, Takaesu G, Zhang W, Kang JS. Close encounters: regulation of vertebrate skeletal myogenesis by cell-cell contact. J Cell Sci. 2005;118:2355–2362. - PubMed
-
- Nagase H, Woessner JF. Matrix metalloproteinases. J Biol Chem. 1999;274:21491–21494. - PubMed
-
- McCawley LJ, Matrisian LM. Matrix metalloproteinases: they’re not just for matrix anymore! Curr Opin Cell Biol. 2001;13:534–540. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Research Materials
Miscellaneous
