

A phosphatase activity of Sts-1 contributes to the suppression of TCR signaling
- PMID: 17679096
- PMCID: PMC2709417
- DOI: 10.1016/j.molcel.2007.06.015
A phosphatase activity of Sts-1 contributes to the suppression of TCR signaling
Abstract
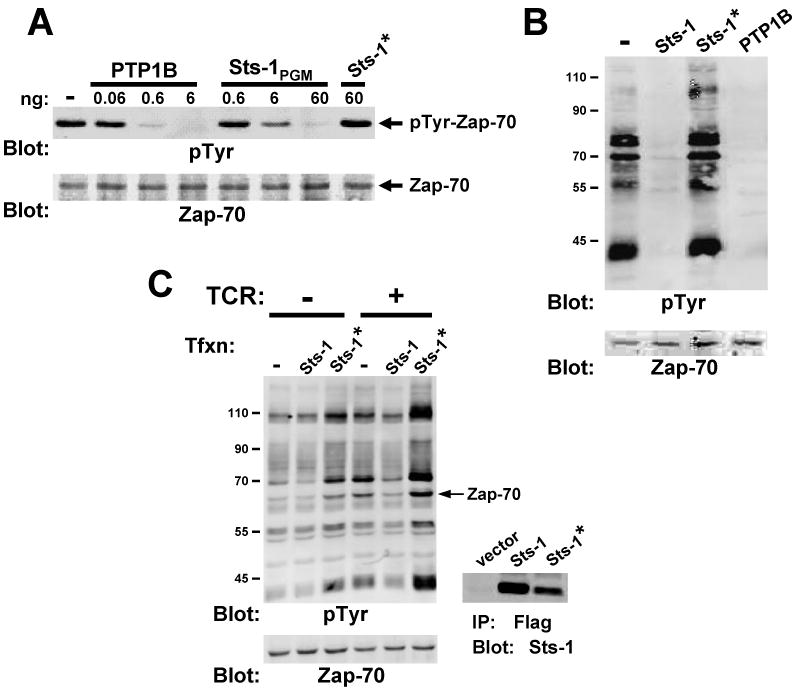
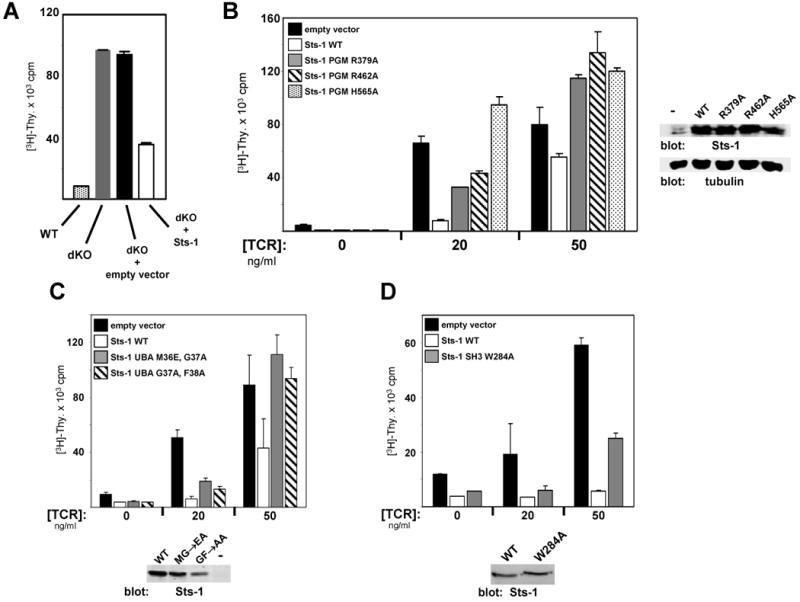
Precise signaling by the T cell receptor (TCR) is crucial for a proper immune response. To ensure that T cells respond appropriately to antigenic stimuli, TCR signaling pathways are subject to multiple levels of regulation. Sts-1 negatively regulates signaling pathways downstream of the TCR by an unknown mechanism(s). Here, we demonstrate that Sts-1 is a phosphatase that can target the tyrosine kinase Zap-70 among other proteins. The X-ray structure of the Sts-1 C terminus reveals that it has homology to members of the phosphoglycerate mutase/acid phosphatase (PGM/AcP) family of enzymes, with residues known to be important for PGM/AcP catalytic activity conserved in nature and position in Sts-1. Point mutations that impair Sts-1 phosphatase activity in vitro also impair the ability of Sts-1 to regulate TCR signaling in T cells. These observations reveal a PGM/AcP-like enzyme activity involved in the control of antigen receptor signaling.
Figures
Similar articles
-
Sts-2 is a phosphatase that negatively regulates zeta-associated protein (ZAP)-70 and T cell receptor signaling pathways.J Biol Chem. 2011 May 6;286(18):15943-54. doi: 10.1074/jbc.M110.177634. Epub 2011 Mar 10. J Biol Chem. 2011. PMID: 21393235 Free PMC article.
-
Insights into the suppressor of T-cell receptor (TCR) signaling-1 (Sts-1)-mediated regulation of TCR signaling through the use of novel substrate-trapping Sts-1 phosphatase variants.FEBS J. 2014 Feb;281(3):696-707. doi: 10.1111/febs.12615. Epub 2013 Dec 12. FEBS J. 2014. PMID: 24256567 Free PMC article.
-
Regulation of ZAP-70 activation and TCR signaling by two related proteins, Sts-1 and Sts-2.Immunity. 2004 Jan;20(1):37-46. doi: 10.1016/s1074-7613(03)00351-0. Immunity. 2004. PMID: 14738763
-
The Sts Proteins: Modulators of Host Immunity.Int J Mol Sci. 2023 May 16;24(10):8834. doi: 10.3390/ijms24108834. Int J Mol Sci. 2023. PMID: 37240179 Free PMC article. Review.
-
TULA-family proteins: Jacks of many trades and then some.J Cell Physiol. 2018 Jan;234(1):274-288. doi: 10.1002/jcp.26890. Epub 2018 Aug 4. J Cell Physiol. 2018. PMID: 30076707 Review.
Cited by
-
Once phosphorylated, tyrosines in carboxyl terminus of protein-tyrosine kinase Syk interact with signaling proteins, including TULA-2, a negative regulator of mast cell degranulation.J Biol Chem. 2012 Mar 9;287(11):8194-204. doi: 10.1074/jbc.M111.326850. Epub 2012 Jan 20. J Biol Chem. 2012. PMID: 22267732 Free PMC article.
-
A novel histidine tyrosine phosphatase, TULA-2, associates with Syk and negatively regulates GPVI signaling in platelets.Blood. 2010 Oct 7;116(14):2570-8. doi: 10.1182/blood-2010-02-268136. Epub 2010 Jun 28. Blood. 2010. PMID: 20585042 Free PMC article.
-
A conserved motif mediates both multimer formation and allosteric activation of phosphoglycerate mutase 5.J Biol Chem. 2014 Sep 5;289(36):25137-48. doi: 10.1074/jbc.M114.565549. Epub 2014 Jul 10. J Biol Chem. 2014. PMID: 25012655 Free PMC article.
-
TULA Proteins in Men, Mice, Hens, and Lice: Welcome to the Family.Int J Mol Sci. 2023 May 23;24(11):9126. doi: 10.3390/ijms24119126. Int J Mol Sci. 2023. PMID: 37298079 Free PMC article. Review.
-
UBASH3A deficiency accelerates type 1 diabetes development and enhances salivary gland inflammation in NOD mice.Sci Rep. 2020 Jul 21;10(1):12019. doi: 10.1038/s41598-020-68956-6. Sci Rep. 2020. PMID: 32694640 Free PMC article.
References
-
- Alonso A, Sasin J, Bottini N, Friedberg I, Friedberg I, Osterman A, Godzik A, Hunter T, Dixon J, Mustelin T. Protein tyrosine phosphatases in the human genome. Cell. 2004;117:699–711. - PubMed
-
- Bond CS, White MF, Hunter WN. High resolution structure of phosphohistidine-activated form of Escherichia co-factor dependent phosphoglycerate mutase. J Biol Chem. 2001;276:3247–3253. - PubMed
-
- Brünger AT. Free R value: a novel statistical quantity for assessing the accuracy of crystal structures. Nature. 1992;355:472–475. - PubMed
Publication types
MeSH terms
Substances
Associated data
- Actions
- Actions
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Research Materials
