

Metabolic stress signaling mediated by mixed-lineage kinases
- PMID: 17679097
- PMCID: PMC1986670
- DOI: 10.1016/j.molcel.2007.07.008
Metabolic stress signaling mediated by mixed-lineage kinases
Abstract
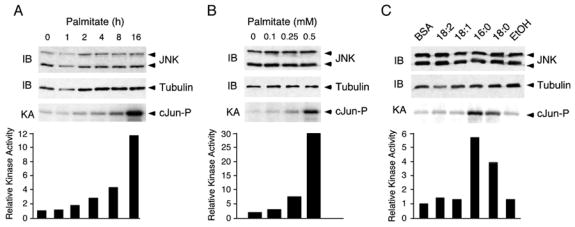
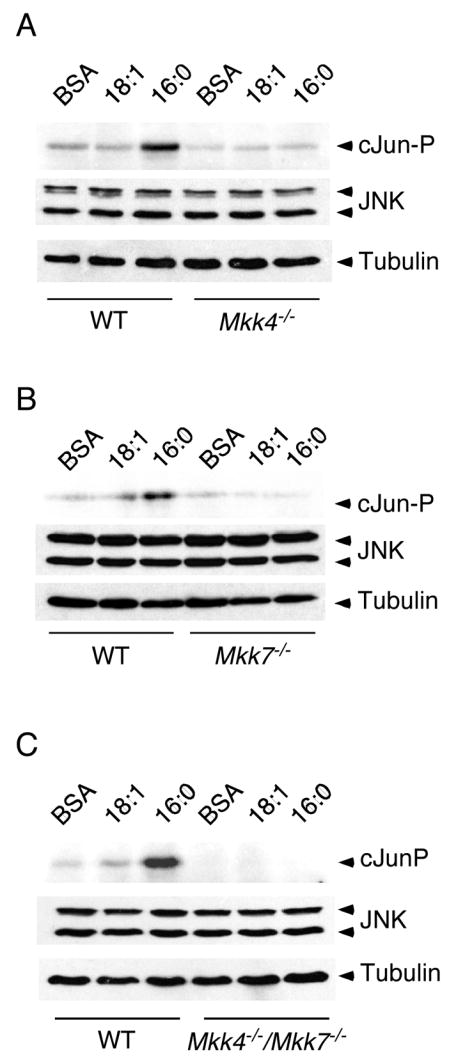
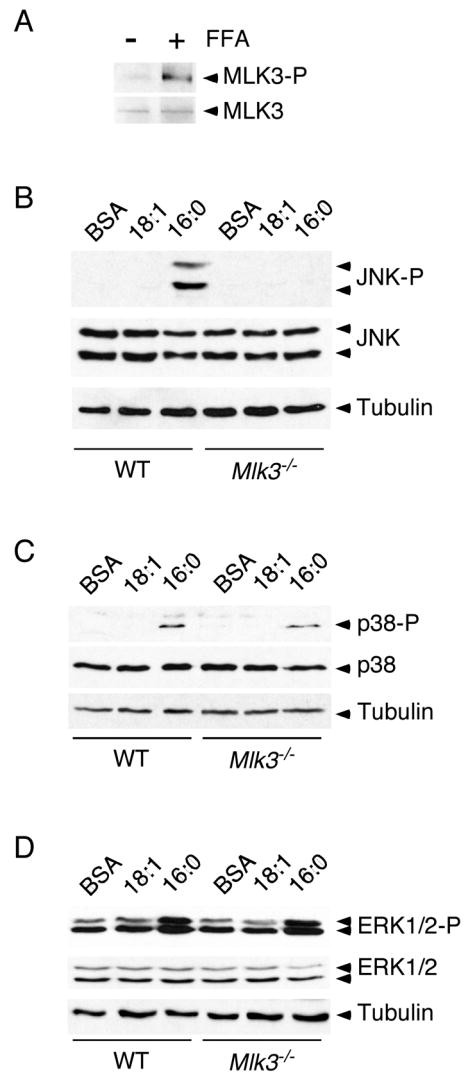
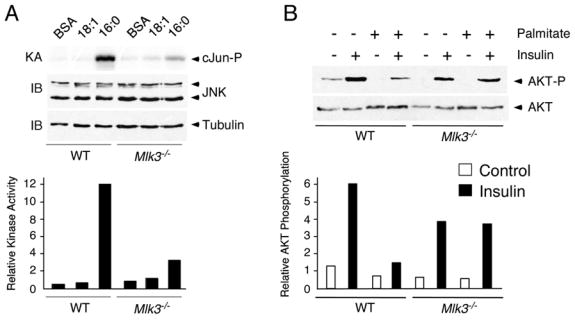
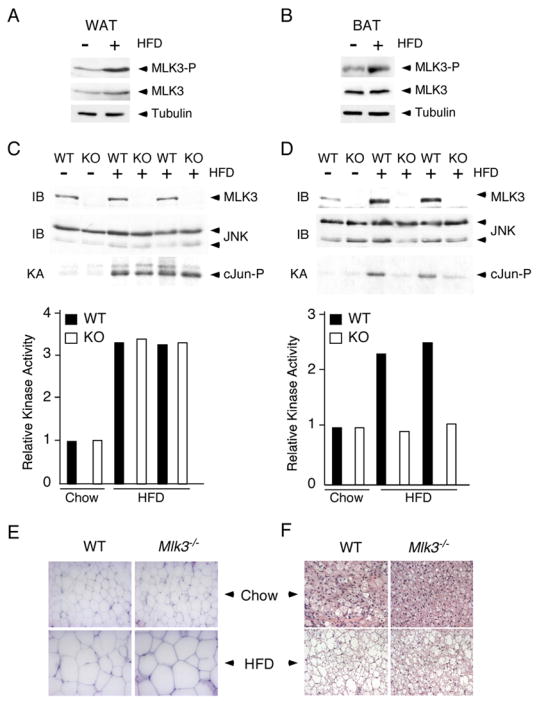
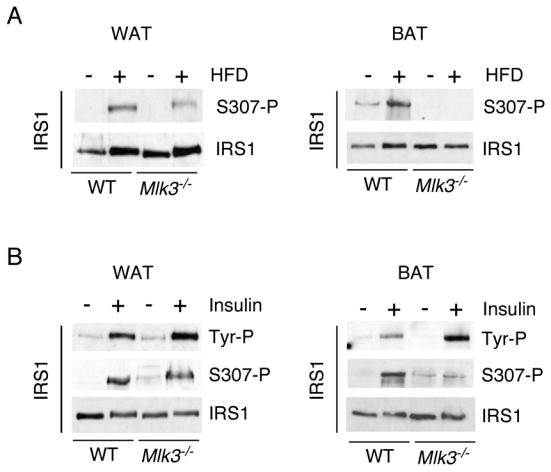
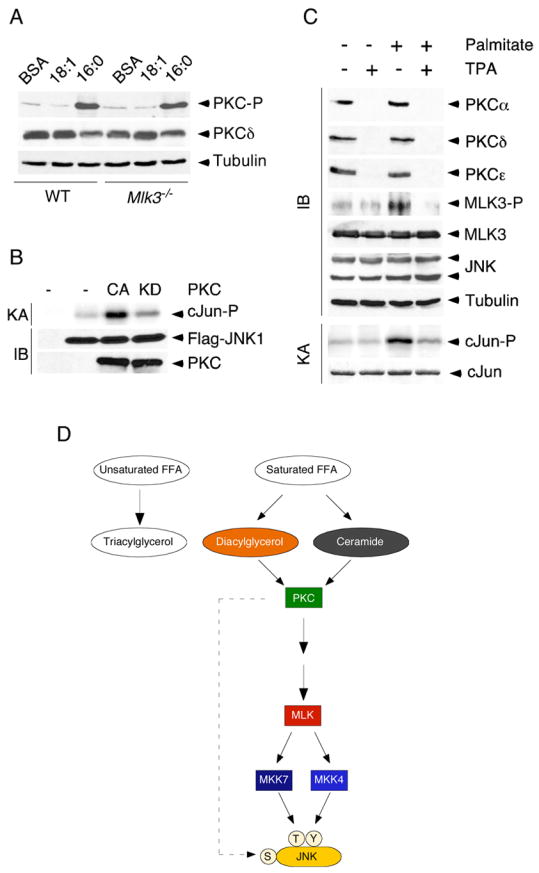
Saturated free fatty acid (FFA) is a major source of metabolic stress that activates the c-Jun NH(2)-terminal kinase (JNK). This FFA-stimulated JNK pathway is relevant to hallmarks of metabolic syndrome, including insulin resistance. Here we used gene ablation studies in mice to demonstrate a central role for mixed-lineage protein kinases (MLK) in this signaling pathway. Saturated FFA causes protein kinase C (PKC)-dependent activation of MLK3 that subsequently causes increased JNK activity by a mechanism that requires the MAP kinase kinases MKK4 and MKK7. Loss of PKC, MLK3, MKK4, or MKK7 expression prevents FFA-stimulated JNK activation. Together, these data establish a signaling pathway that mediates effects of metabolic stress on insulin resistance.
Figures
References
-
- Aguirre V, Uchida T, Yenush L, Davis R, White MF. The c-Jun NH(2)-terminal kinase promotes insulin resistance during association with insulin receptor substrate-1 and phosphorylation of Ser(307) J Biol Chem. 2000;275:9047–9054. - PubMed
-
- Aguirre V, Werner ED, Giraud J, Lee YH, Shoelson SE, White MF. Phosphorylation of Ser307 in insulin receptor substrate-1 blocks interactions with the insulin receptor and inhibits insulin action. J Biol Chem. 2002;277:1531–1537. - PubMed
-
- Arner P. Insulin resistance in type 2 diabetes: role of fatty acids. Diabetes Metab Res Rev. 2002;18(Suppl 2):S5–9. - PubMed
-
- Barthwal MK, Sathyanarayana P, Kundu CN, Rana B, Pradeep A, Sharma C, Woodgett JR, Rana A. Negative regulation of mixed lineage kinase 3 by protein kinase B/AKT leads to cell survival. J Biol Chem. 2003;278:3897–3902. - PubMed
-
- Boden G. Effects of free fatty acids (FFA) on glucose metabolism: significance for insulin resistance and type 2 diabetes. Exp Clin Endocrinol Diabetes. 2003;111:121–124. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Research Materials
Miscellaneous
