

The intriguing normal acute inflammatory response in mice lacking vimentin
- PMID: 17680824
- PMCID: PMC2219279
- DOI: 10.1111/j.1365-2249.2007.03460.x
The intriguing normal acute inflammatory response in mice lacking vimentin
Abstract
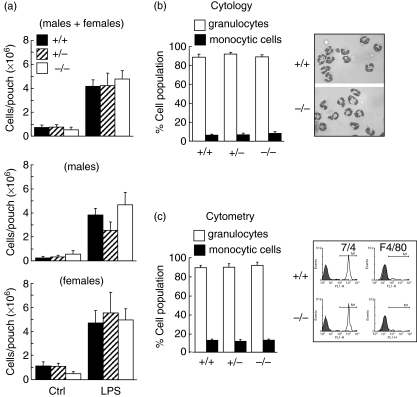
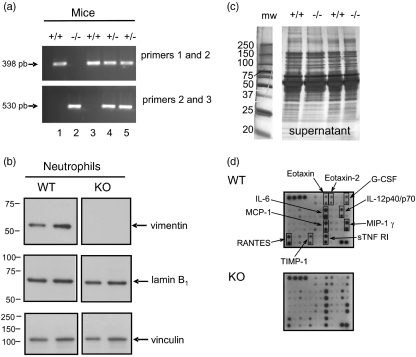
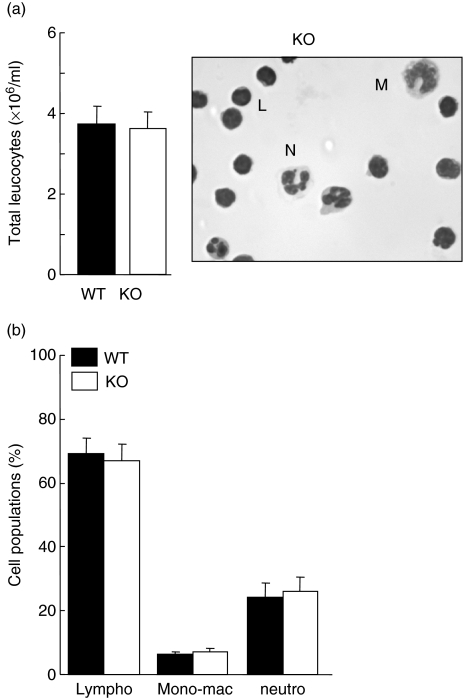
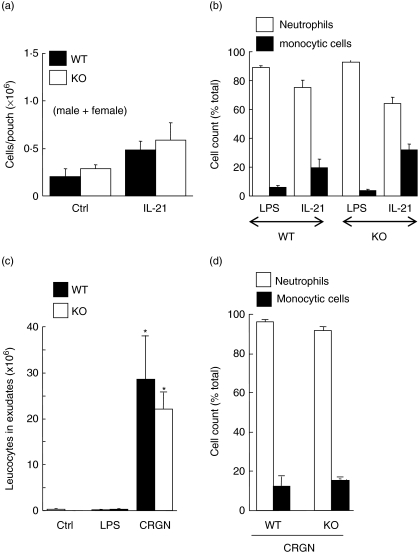
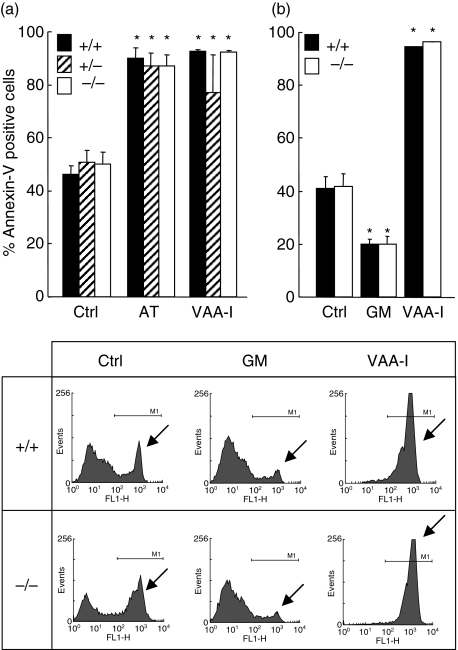
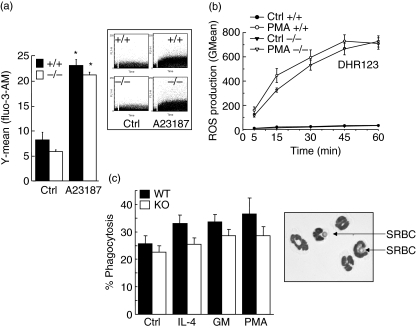
Neutrophils express only two intermediate filament proteins, vimentin and, to a lesser extent, lamin B. Lamin B mutant mice die shortly after birth; however, mice lacking vimentin (vim(-/-)) develop and reproduce normally. Herein, we investigate for the first time the role of vimentin in general inflammation in vivo and in neutrophil functions ex vivo. Using the murine air pouch model, we show that the inflammatory response induced by lipopolysaccharide, interleukin-21 or carageenan is, intriguingly, uncompromised in vim(-/-) mice and that neutrophil functions are not altered ex vivo. Our results suggest that vimentin is dispensable for the establishment of an acute inflammatory response in vivo. In addition, based on several criteria presented in this study, one has to accept the existence of a very complex compensatory mechanism to explain the intriguing normal inflammatory response in absence of vimentin.
Figures
Similar articles
-
Cell surface expression of intermediate filament proteins vimentin and lamin B1 in human neutrophil spontaneous apoptosis.J Leukoc Biol. 2006 Mar;79(3):489-98. doi: 10.1189/jlb.0405190. Epub 2005 Dec 19. J Leukoc Biol. 2006. PMID: 16365157
-
Impaired flow-induced dilation in mesenteric resistance arteries from mice lacking vimentin.J Clin Invest. 1997 Dec 1;100(11):2909-14. doi: 10.1172/JCI119840. J Clin Invest. 1997. PMID: 9389758 Free PMC article.
-
Spontaneously immortalized mouse embryo fibroblasts: growth behavior of wild-type and vimentin-deficient cells in relation to mitochondrial structure and activity.DNA Cell Biol. 2005 Nov;24(11):680-709. doi: 10.1089/dna.2005.24.680. DNA Cell Biol. 2005. PMID: 16274292
-
Vimentin: the conundrum of the intermediate filament gene family.Bioessays. 1998 Jan;20(1):79-86. doi: 10.1002/(SICI)1521-1878(199801)20:1<79::AID-BIES11>3.0.CO;2-5. Bioessays. 1998. PMID: 9504050 Review.
-
Redox regulation of neutrophil apoptosis and the systemic inflammatory response syndrome.Clin Sci (Lond). 2005 May;108(5):413-24. doi: 10.1042/CS20040228. Clin Sci (Lond). 2005. PMID: 15831090 Review.
Cited by
-
Downregulation of vimentin in macrophages infected with live Mycobacterium tuberculosis is mediated by Reactive Oxygen Species.Sci Rep. 2016 Feb 15;6:21526. doi: 10.1038/srep21526. Sci Rep. 2016. PMID: 26876331 Free PMC article.
-
Senescent cells and macrophages: key players for regeneration?Open Biol. 2020 Dec;10(12):200309. doi: 10.1098/rsob.200309. Epub 2020 Dec 23. Open Biol. 2020. PMID: 33352064 Free PMC article. Review.
-
The regulatory role of cell mechanics for migration of differentiating myeloid cells.Proc Natl Acad Sci U S A. 2009 Sep 15;106(37):15696-701. doi: 10.1073/pnas.0811261106. Epub 2009 Aug 26. Proc Natl Acad Sci U S A. 2009. PMID: 19717452 Free PMC article.
-
Single-cell transcriptome analysis of cavernous tissues reveals the key roles of pericytes in diabetic erectile dysfunction.Elife. 2024 Jun 10;12:RP88942. doi: 10.7554/eLife.88942. Elife. 2024. PMID: 38856719 Free PMC article.
-
The LINC-less granulocyte nucleus.Eur J Cell Biol. 2009 Apr;88(4):203-14. doi: 10.1016/j.ejcb.2008.10.001. Epub 2008 Nov 18. Eur J Cell Biol. 2009. PMID: 19019491 Free PMC article.
References
-
- Bruel A, Paschke S, Jainta S, et al. Remodeling of vimentin cytoskeleton correlates with enhanced motility of promyelocytic leukemia cells during differentiation induced by retinoic acid. Anticancer Res. 2001;21:3973–80. - PubMed
-
- Colucci-Guyon E, Portier MM, Dunia I, Paulin D, Pournin S, Babinet C. Mice lacking vimentin develop and reproduce without an obvious phenotype. Cell. 1994;79:679–94. - PubMed
-
- Schiffers PM, Henrion D, Boulanger CM, et al. Altered flow-induced arterial remodeling in vimentin-deficient mice. Arterioscler Thromb Vasc Biol. 2000;20:611–6. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
Miscellaneous
