

Regulated HsSAS-6 levels ensure formation of a single procentriole per centriole during the centrosome duplication cycle
- PMID: 17681132
- PMCID: PMC2628752
- DOI: 10.1016/j.devcel.2007.07.004
Regulated HsSAS-6 levels ensure formation of a single procentriole per centriole during the centrosome duplication cycle
Abstract
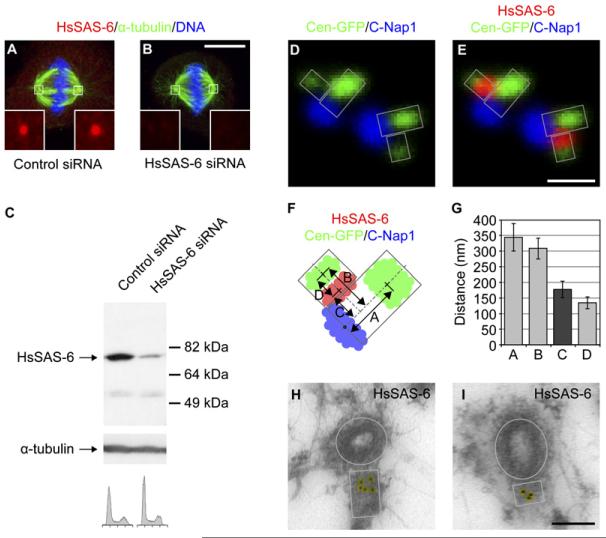
Centrosome duplication involves the formation of a single procentriole next to each centriole, once per cell cycle. The mechanisms governing procentriole formation and those restricting its occurrence to one event per centriole are poorly understood. Here, we show that HsSAS-6 is necessary for procentriole formation and that it localizes asymmetrically next to the centriole at the onset of procentriole formation. HsSAS-6 levels oscillate during the cell cycle, with the protein being degraded in mitosis and starting to accumulate again at the end of the following G1. Our findings indicate that APC(Cdh1) targets HsSAS-6 for degradation by the 26S proteasome. Importantly, we demonstrate that increased HsSAS-6 levels promote formation of more than one procentriole per centriole. Therefore, regulated HsSAS-6 levels normally ensure that each centriole seeds the formation of a single procentriole per cell cycle, thus playing a fundamental role in driving the centrosome duplication cycle and ensuring genome integrity.
Figures






References
-
- Byers B, Goetsch L. Duplication of spindle plaques and integration of the yeast cell cycle. Cold Spring Harb. Symp. Quant. Biol. 1974;38:123–131. - PubMed
-
- Chretien D, Buendia B, Fuller SD, Karsenti E. Reconstruction of the centrosome cycle from cryoelectron micrographs. J. Struct. Biol. 1997;120:117–133. - PubMed
-
- Dammermann A, Muller-Reichert T, Pelletier L, Habermann B, Desai A, Oegema K. Centriole assembly requires both centriolar and pericentriolar material proteins. Dev. Cell. 2004;7:815–829. - PubMed
-
- Euteneuer U, Graf R, Kube-Granderath E, Schliwa M. Dictyostelium gamma-tubulin: molecular characterization and ultra-structural localization. J. Cell Sci. 1998;111:405–412. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Miscellaneous
