

DNA topoisomerase II, genotoxicity, and cancer
- PMID: 17681352
- PMCID: PMC2679583
- DOI: 10.1016/j.mrfmmm.2007.06.009
DNA topoisomerase II, genotoxicity, and cancer
Abstract
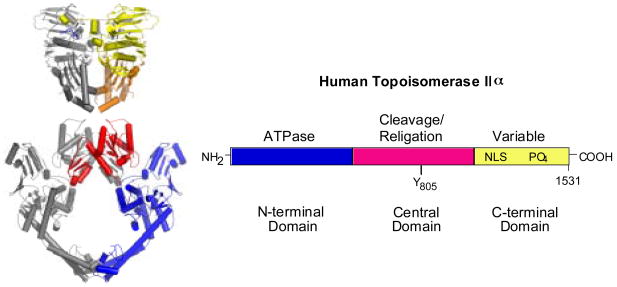
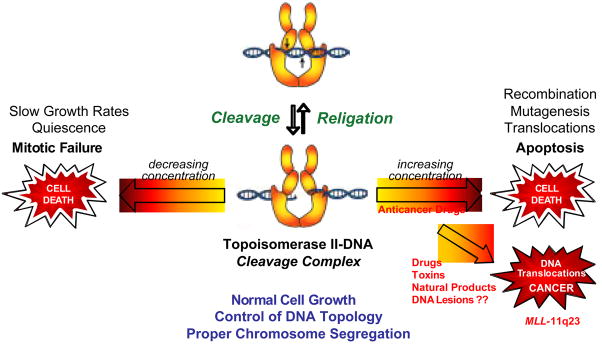
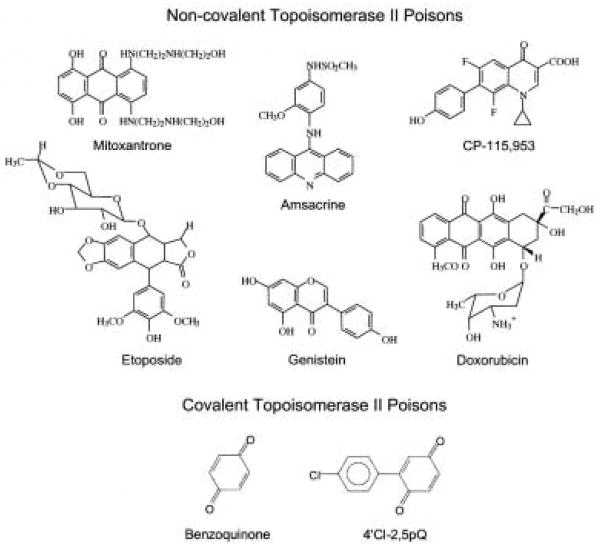
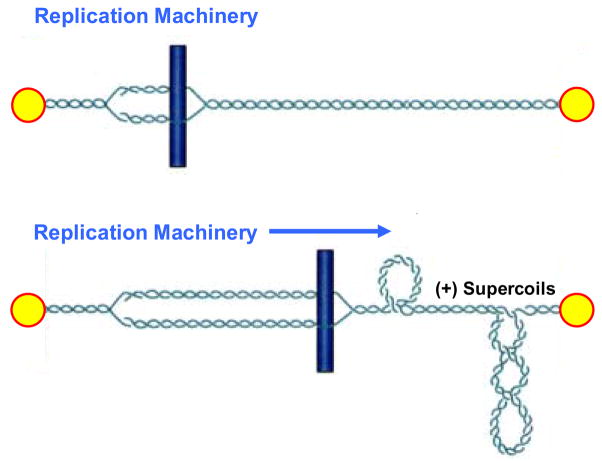
Type II topoisomerases are ubiquitous enzymes that play essential roles in a number of fundamental DNA processes. They regulate DNA under- and overwinding, and resolve knots and tangles in the genetic material by passing an intact double helix through a transient double-stranded break that they generate in a separate segment of DNA. Because type II topoisomerases generate DNA strand breaks as a requisite intermediate in their catalytic cycle, they have the potential to fragment the genome every time they function. Thus, while these enzymes are essential to the survival of proliferating cells, they also have significant genotoxic effects. This latter aspect of type II topoisomerase has been exploited for the development of several classes of anticancer drugs that are widely employed for the clinical treatment of human malignancies. However, considerable evidence indicates that these enzymes also trigger specific leukemic chromosomal translocations. In light of the impact, both positive and negative, of type II topoisomerases on human cells, it is important to understand how these enzymes function and how their actions can destabilize the genome. This article discusses both aspects of human type II topoisomerases.
Figures
References
-
- Watson JD, Crick FHC. Molecular Structure of Nucleic Acids. Nature. 1953;171:737–738. - PubMed
-
- Watson JD, Crick FHC. Genetical Implications of the Structure of Deoxyribonucleic Acid. Nature. 1953;171:964–967. - PubMed
-
- Cozzarelli NR, Wang JC, editors. DNA Topology and its Biological Effects. Cold Spring Harbor Laboratory Press: Cold Spring Harbor; 1990.
-
- Kanaar R, Cozzarelli NR. Roles of supercoiled DNA structure in DNA transactions. Curr Opin Struct Biol. 1992;2:369–379.
-
- Wang JC. DNA topoisomerases. Annu Rev Biochem. 1996;65:635–692. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
