

p32 is a novel mammalian Lgl binding protein that enhances the activity of protein kinase Czeta and regulates cell polarity
- PMID: 17682048
- PMCID: PMC2064465
- DOI: 10.1083/jcb.200612022
p32 is a novel mammalian Lgl binding protein that enhances the activity of protein kinase Czeta and regulates cell polarity
Abstract
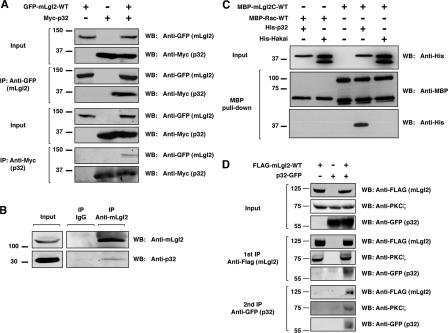
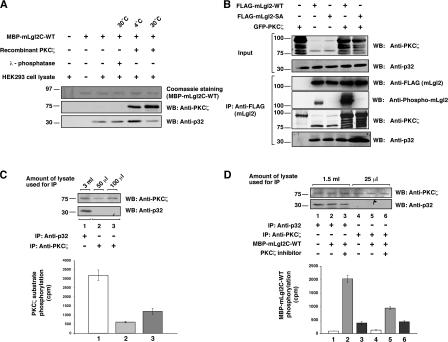
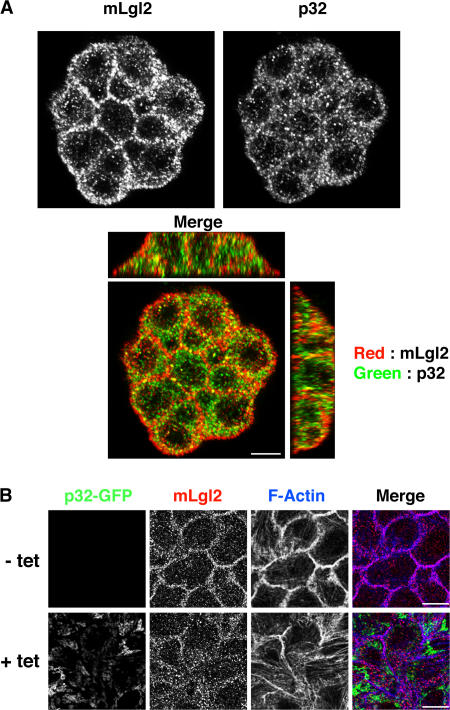
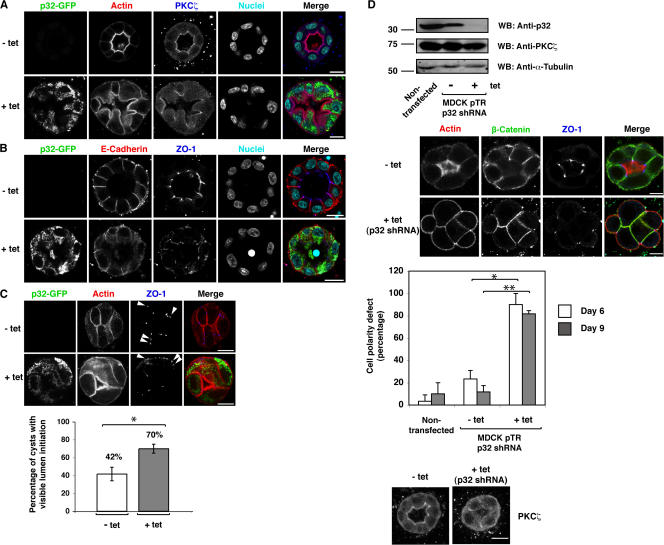
Lgl (lethal giant larvae) plays an important role in cell polarity. Atypical protein kinase C (aPKC) binds to and phosphorylates Lgl, and the phosphorylation negatively regulates Lgl activity. In this study, we identify p32 as a novel Lgl binding protein that directly binds to a domain on mammalian Lgl2 (mLgl2), which contains the aPKC phosphorylation site. p32 also binds to PKCzeta, and the three proteins form a transient ternary complex. When p32 is bound, PKCzeta is stimulated to phosphorylate mLgl2 more efficiently. p32 overexpression in Madin-Darby canine kidney cells cultured in a 3D matrix induces an expansion of the actin-enriched apical membrane domain and disrupts cell polarity. Addition of PKCzeta inhibitor blocks apical actin accumulation, which is rescued by p32 overexpression. p32 knockdown by short hairpin RNA also induces cell polarity defects. Collectively, our data indicate that p32 is a novel regulator of cell polarity that forms a complex with mLgl2 and aPKC and enhances aPKC activity.
Figures
References
-
- Betschinger, J., K. Mechtler, and J.A. Knoblich. 2003. The Par complex directs asymmetric cell division by phosphorylating the cytoskeletal protein Lgl. Nature. 422:326–330. - PubMed
-
- Betschinger, J., F. Eisenhaber, and J.A. Knoblich. 2005. Phosphorylation- induced autoinhibition regulates the cytoskeletal protein Lethal (2) giant larvae. Curr. Biol. 15:276–282. - PubMed
-
- Bilder, D. 2004. Epithelial polarity and proliferation control: links from the Drosophila neoplastic tumor suppressors. Genes Dev. 18:1909–1925. - PubMed
-
- Bilder, D., M. Li, and N. Perrimon. 2000. Cooperative regulation of cell polarity and growth by Drosophila tumor suppressors. Science. 289:113–116. - PubMed
-
- Chalmers, A.D., M. Pambos, J. Mason, S. Lang, C. Wylie, and N. Papalopulu. 2005. aPKC, Crumbs3 and Lgl2 control apicobasal polarity in early vertebrate development. Development. 132:977–986. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Research Materials
