

Polysialic acid-directed migration and differentiation of neural precursors are essential for mouse brain development
- PMID: 17682066
- PMCID: PMC2099222
- DOI: 10.1128/MCB.00205-07
Polysialic acid-directed migration and differentiation of neural precursors are essential for mouse brain development
Abstract
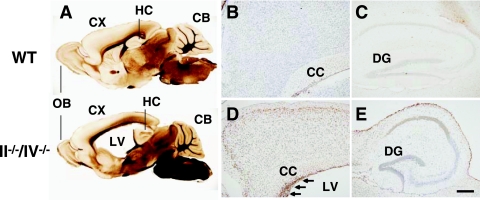
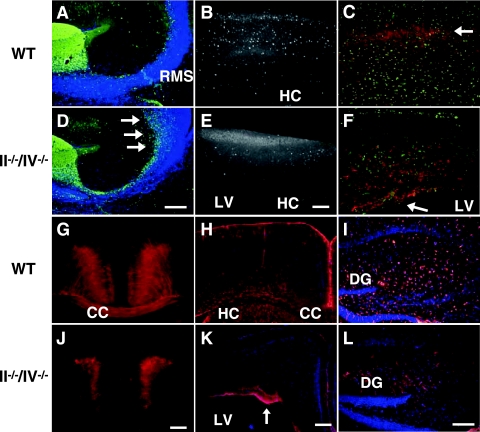
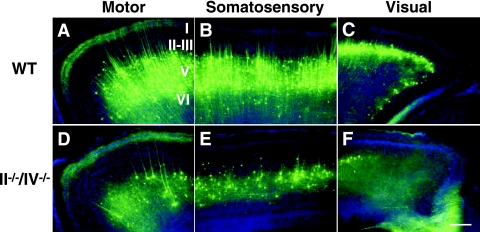
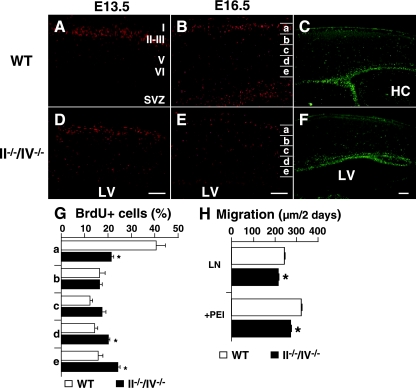
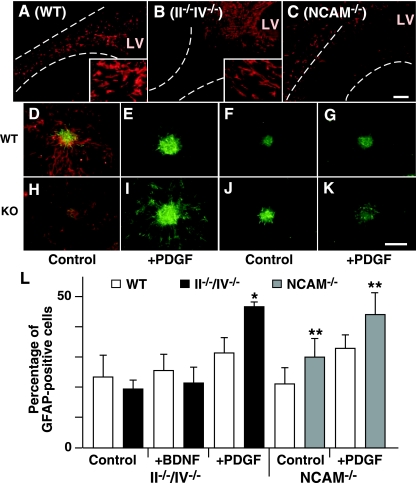
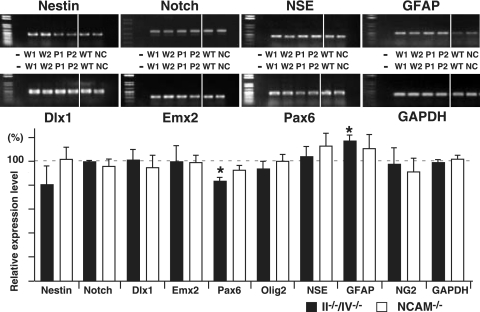
Polysialic acid, which is synthesized by two polysialyltransferases, ST8SiaII and ST8SiaIV, plays an essential role in brain development by modifying the neural cell adhesion molecule (NCAM). It is currently unclear how polysialic acid functions in different processes of neural development. Here we generated mice doubly mutant in both ST8SiaII and ST8SiaIV to determine the effects of loss of polysialic acid on brain development. In contrast to NCAM-deficient, ST8SiaII-deficient, or ST8SiaIV-deficient single mutant mice, ST8SiaII and ST8SiaIV double mutants displayed severe defects in anatomical organization of the forebrain associated with apoptotic cell death. Loss of polysialic acid affected both tangential and radial migration of neural precursors during cortical development, resulting in aberrant positioning of neuronal and glial cells. Glial cell differentiation was aberrantly increased in vivo and in vitro in the absence of polysialic acid. Consistent with these findings, polysialic acid-deficient mice exhibited increased expression of the glial cell marker glial fibrillary acidic protein and a decrease in expression of Pax6, a transcription factor regulating neural cell migration. These results indicate that polysialic acid regulates cell migration and differentiation of neural precursors crucial for brain development.
Figures
References
-
- Angata, K., D. Chan, J. Thibault, and M. Fukuda. 2004. Molecular dissection of the ST8Sia IV polysialyltransferase. Distinct domains are required for neural cell adhesion molecule recognition and polysialylation. J. Biol. Chem. 279:25883-25890. - PubMed
-
- Angata, K., and M. Fukuda. 2003. Polysialyltransferases: major players in polysialic acid synthesis on the neural cell adhesion molecule. Biochimie 85:195-206. - PubMed
-
- Angata, K., J. M. Long, O. Bukalo, W. Lee, A. Dityatev, A. Wynshaw-Boris, M. Schachner, M. Fukuda, and J. D. Marth. 2004. Sialyltransferase ST8Sia-II assembles a subset of polysialic acid that directs hippocampal axonal targeting and promotes fear behavior. J. Biol. Chem. 279:32603-32613. - PubMed
-
- Berglund, E. O., K. K. Murai, B. Fredette, G. Sekerkova, B. Marturano, L. Weber, E. Mugnaini, and B. Ranscht. 1999. Ataxia and abnormal cerebellar microorganization in mice with ablated contactin gene expression. Neuron 24:739-750. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Molecular Biology Databases
Research Materials
Miscellaneous