

Nuclear factors are involved in hepatitis C virus RNA replication
- PMID: 17684232
- PMCID: PMC1986813
- DOI: 10.1261/rna.594207
Nuclear factors are involved in hepatitis C virus RNA replication
Abstract
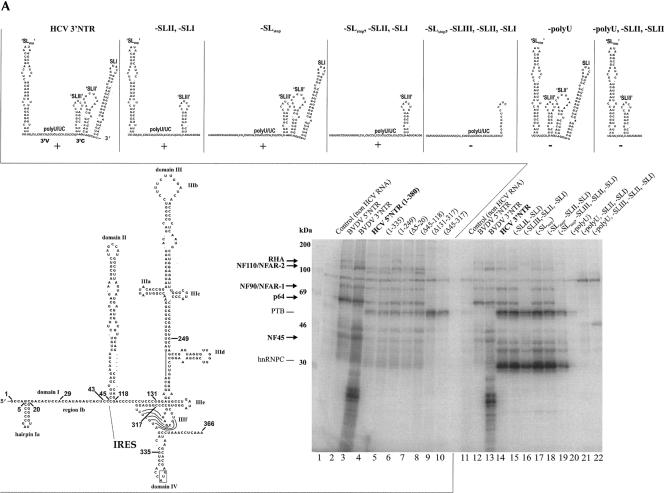
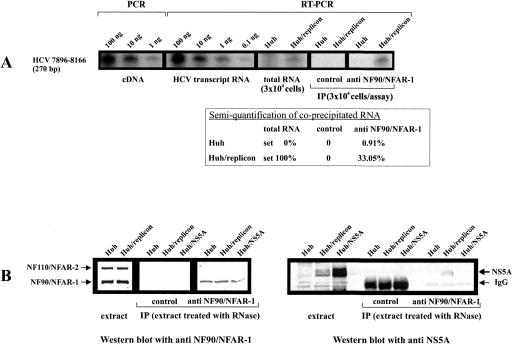
Unraveling the molecular basis of the life cycle of hepatitis C virus (HCV), a prevalent agent of human liver disease, entails the identification of cell-encoded factors that participate in the replication of the viral RNA genome. This study provides evidence that the so-called NF/NFAR proteins, namely, NF90/NFAR-1, NF110/NFAR-2, NF45, and RNA helicase A (RHA), which mostly belong to the dsRBM protein family, are involved in the HCV RNA replication process. NF/NFAR proteins were shown to specifically bind to replication signals in the HCV genomic 5' and 3' termini and to promote the formation of a looplike structure of the viral RNA. In cells containing replicating HCV RNA, the generally nuclear NF/NFAR proteins accumulate in the cytoplasmic viral replication complexes, and the prototype NFAR protein, NF90/NFAR-1, stably interacts with a viral protein. HCV replication was inhibited in cells where RNAi depleted RHA from the cytoplasm. Likewise, HCV replication was hindered in cells that contained another NF/NFAR protein recruiting virus. The recruitment of NF/NFAR proteins by HCV is assumed to serve two major purposes: to support 5'-3' interactions of the viral RNA for the coordination of viral protein and RNA synthesis and to weaken host-defense mechanisms.
Figures








References
-
- Behrens, S.E., Isken, O. Cis and trans-acting factors in Flaviviridae replication. In: Kalitzky M., Borowski P., editors. Molecular Biology of the Flavivirus. Horizon Bioscience; Norwich, UK: 2006. pp. 101–134.
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Miscellaneous