

Molecular components and functions of the endocannabinoid system in mouse prefrontal cortex
- PMID: 17684555
- PMCID: PMC1933592
- DOI: 10.1371/journal.pone.0000709
Molecular components and functions of the endocannabinoid system in mouse prefrontal cortex
Abstract
Background: Cannabinoids have deleterious effects on prefrontal cortex (PFC)-mediated functions and multiple evidences link the endogenous cannabinoid (endocannabinoid) system, cannabis use and schizophrenia, a disease in which PFC functions are altered. Nonetheless, the molecular composition and the physiological functions of the endocannabinoid system in the PFC are unknown.
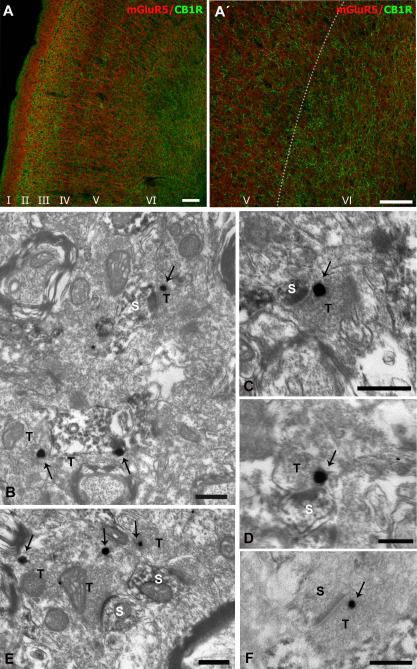
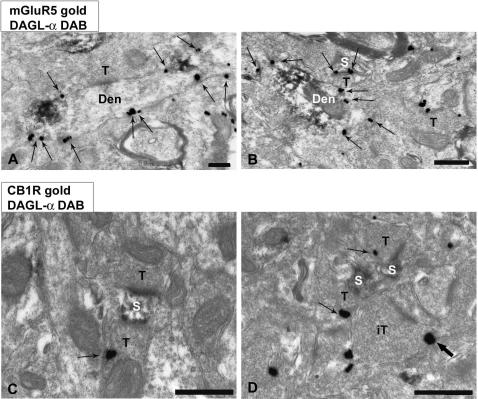
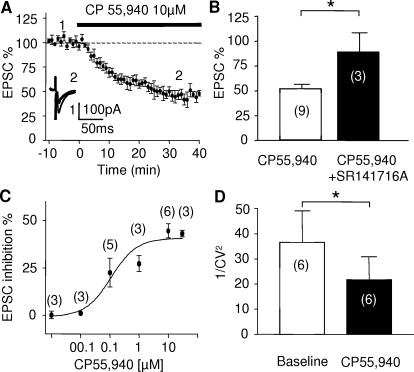
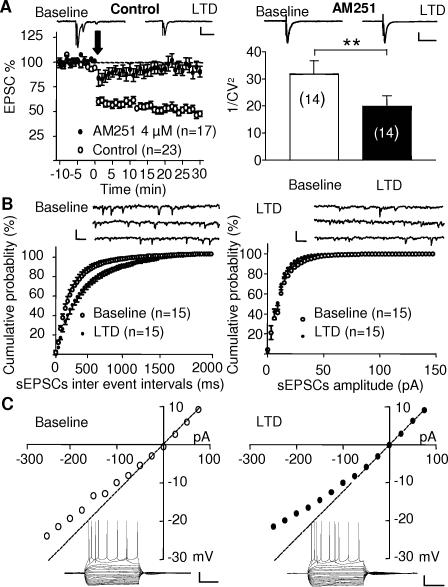
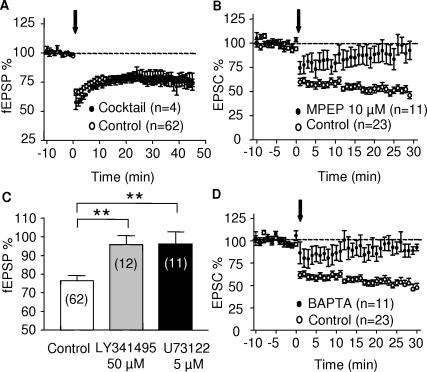
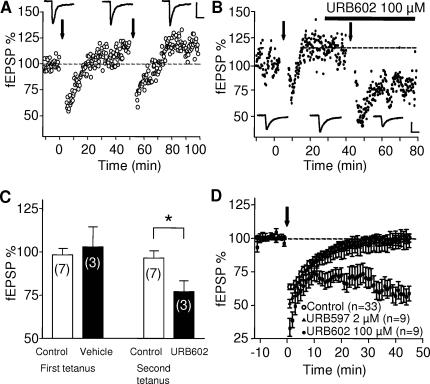
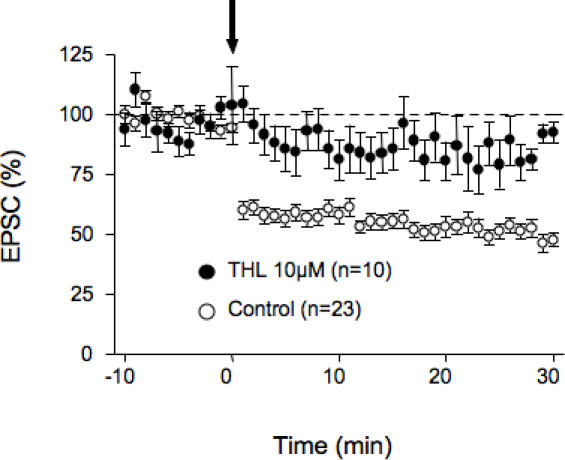
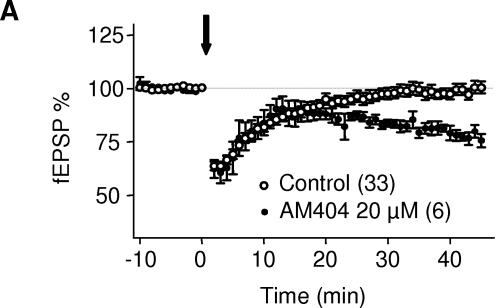
Methodology/principal findings: Here, using electron microscopy we found that key proteins involved in endocannabinoid signaling are expressed in layers v/vi of the mouse prelimbic area of the PFC: presynaptic cannabinoid CB1 receptors (CB1R) faced postsynaptic mGluR5 while diacylglycerol lipase alpha (DGL-alpha), the enzyme generating the endocannabinoid 2-arachidonoyl-glycerol (2-AG) was expressed in the same dendritic processes as mGluR5. Activation of presynaptic CB1R strongly inhibited evoked excitatory post-synaptic currents. Prolonged synaptic stimulation at 10Hz induced a profound long-term depression (LTD) of layers V/VI excitatory inputs. The endocannabinoid -LTD was presynaptically expressed and depended on the activation of postsynaptic mGluR5, phospholipase C and a rise in postsynaptic Ca(2+) as predicted from the localization of the different components of the endocannabinoid system. Blocking the degradation of 2-AG (with URB 602) but not of anandamide (with URB 597) converted subthreshold tetanus to LTD-inducing ones. Moreover, inhibiting the synthesis of 2-AG with Tetrahydrolipstatin, blocked endocannabinoid-mediated LTD. All together, our data show that 2-AG mediates LTD at these synapses.
Conclusions/significance: Our data show that the endocannabinoid -retrograde signaling plays a prominent role in long-term synaptic plasticity at the excitatory synapses of the PFC. Alterations of endocannabinoid -mediated synaptic plasticity may participate to the etiology of PFC-related pathologies.
Conflict of interest statement
Figures
Similar articles
-
Molecular architecture of endocannabinoid signaling at nociceptive synapses mediating analgesia.Eur J Neurosci. 2009 May;29(10):1964-78. doi: 10.1111/j.1460-9568.2009.06751.x. Epub 2009 May 9. Eur J Neurosci. 2009. PMID: 19453631 Free PMC article.
-
Activation of type 5 metabotropic glutamate receptors and diacylglycerol lipase-α initiates 2-arachidonoylglycerol formation and endocannabinoid-mediated analgesia.J Neurosci. 2012 Jul 11;32(28):9457-68. doi: 10.1523/JNEUROSCI.0013-12.2012. J Neurosci. 2012. PMID: 22787031 Free PMC article.
-
Postsynaptic diacylglycerol lipase mediates retrograde endocannabinoid suppression of inhibition in mouse prefrontal cortex.J Physiol. 2011 Oct 15;589(Pt 20):4857-84. doi: 10.1113/jphysiol.2011.212225. Epub 2011 Aug 1. J Physiol. 2011. PMID: 21807615 Free PMC article.
-
Biosynthesis and degradation of the endocannabinoid 2-arachidonoylglycerol.Biofactors. 2011 Jan-Feb;37(1):1-7. doi: 10.1002/biof.131. Epub 2010 Nov 29. Biofactors. 2011. PMID: 21328621 Review.
-
Endocannabinoid-mediated retrograde modulation of synaptic transmission.Curr Opin Neurobiol. 2014 Dec;29:1-8. doi: 10.1016/j.conb.2014.03.017. Epub 2014 Apr 18. Curr Opin Neurobiol. 2014. PMID: 24747340 Review.
Cited by
-
The role of dopamine and endocannabinoid systems in prefrontal cortex development: Adolescence as a critical period.Front Neural Circuits. 2022 Nov 1;16:939235. doi: 10.3389/fncir.2022.939235. eCollection 2022. Front Neural Circuits. 2022. PMID: 36389180 Free PMC article. Review.
-
Perinatal THC exposure via lactation induces lasting alterations to social behavior and prefrontal cortex function in rats at adulthood.Neuropsychopharmacology. 2020 Oct;45(11):1826-1833. doi: 10.1038/s41386-020-0716-x. Epub 2020 May 19. Neuropsychopharmacology. 2020. PMID: 32428929 Free PMC article.
-
Endocannabinoid modulation of cortical up-states and NREM sleep.PLoS One. 2014 Feb 10;9(2):e88672. doi: 10.1371/journal.pone.0088672. eCollection 2014. PLoS One. 2014. PMID: 24520411 Free PMC article.
-
Cannabinoids and Vanilloids in Schizophrenia: Neurophysiological Evidence and Directions for Basic Research.Front Pharmacol. 2017 Jun 21;8:399. doi: 10.3389/fphar.2017.00399. eCollection 2017. Front Pharmacol. 2017. PMID: 28680405 Free PMC article. Review.
-
Molecular architecture of endocannabinoid signaling at nociceptive synapses mediating analgesia.Eur J Neurosci. 2009 May;29(10):1964-78. doi: 10.1111/j.1460-9568.2009.06751.x. Epub 2009 May 9. Eur J Neurosci. 2009. PMID: 19453631 Free PMC article.
References
-
- Chevaleyre V, Takahashi KA, Castillo PE. Endocannabinoid-mediated synaptic plasticity in the CNS. Annu Rev Neurosci. 2006;29:37–76. - PubMed
-
- Fride E. Endocannabinoids in the central nervous system: from neuronal networks to behavior. Curr Drug Targets CNS Neurol Disord. 2005;4:633–642. - PubMed
-
- Howlett AC. The cannabinoid receptors. Prostaglandins Other Lipid Mediat. 2002;68-69:619–631. - PubMed
-
- Mackie K. Cannabinoid receptors as therapeutic targets. Annu Rev Pharmacol Toxicol. 2006;46:101–122. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
Miscellaneous
