

Plasticity of the intrinsic period of the human circadian timing system
- PMID: 17684566
- PMCID: PMC1934931
- DOI: 10.1371/journal.pone.0000721
Plasticity of the intrinsic period of the human circadian timing system
Abstract
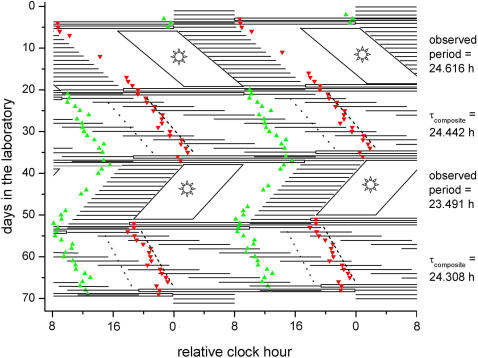
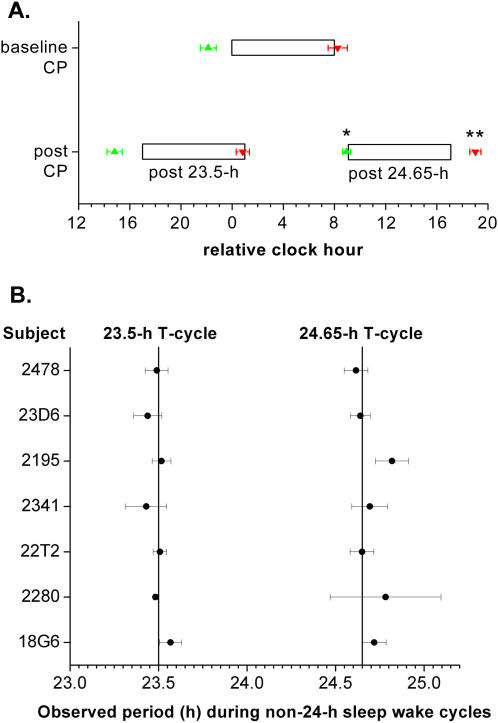
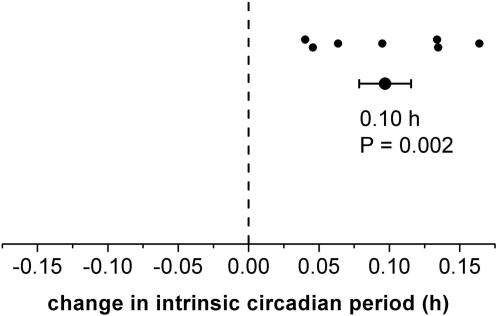
Human expeditions to Mars will require adaptation to the 24.65-h Martian solar day-night cycle (sol), which is outside the range of entrainment of the human circadian pacemaker under lighting intensities to which astronauts are typically exposed. Failure to entrain the circadian time-keeping system to the desired rest-activity cycle disturbs sleep and impairs cognitive function. Furthermore, differences between the intrinsic circadian period and Earth's 24-h light-dark cycle underlie human circadian rhythm sleep disorders, such as advanced sleep phase disorder and non-24-hour sleep-wake disorders. Therefore, first, we tested whether exposure to a model-based lighting regimen would entrain the human circadian pacemaker at a normal phase angle to the 24.65-h Martian sol and to the 23.5-h day length often required of astronauts during short duration space exploration. Second, we tested here whether such prior entrainment to non-24-h light-dark cycles would lead to subsequent modification of the intrinsic period of the human circadian timing system. Here we show that exposure to moderately bright light ( approximately 450 lux; approximately 1.2 W/m(2)) for the second or first half of the scheduled wake episode is effective for entraining individuals to the 24.65-h Martian sol and a 23.5-h day length, respectively. Estimations of the circadian periods of plasma melatonin, plasma cortisol, and core body temperature rhythms collected under forced desynchrony protocols revealed that the intrinsic circadian period of the human circadian pacemaker was significantly longer following entrainment to the Martian sol as compared to following entrainment to the 23.5-h day. The latter finding of after-effects of entrainment reveals for the first time plasticity of the period of the human circadian timing system. Both findings have important implications for the treatment of circadian rhythm sleep disorders and human space exploration.
Conflict of interest statement
Figures
References
-
- Borbély AA. A two process model of sleep regulation. Hum Neurobiol. 1982;1:195–204. - PubMed
-
- Dijk DJ, Czeisler CA. Paradoxical timing of the circadian rhythm of sleep propensity serves to consolidate sleep and wakefulness in humans. Neurosci Lett. 1994;166:63–68. - PubMed
-
- Ibuka N, Kawamura H. Loss of circadian rhythm in sleep-wakefulness cycle in the rat by suprachiasmatic nucleus lesions. Brain Res. 1975;96:76–81. - PubMed
-
- Boivin DB, Duffy JF, Kronauer RE, Czeisler CA. Dose-response relationships for resetting of human circadian clock by light. Nature. 1996;379:540–542. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Miscellaneous
