

Complement induction in spinal cord microglia results in anaphylatoxin C5a-mediated pain hypersensitivity
- PMID: 17687047
- PMCID: PMC6672952
- DOI: 10.1523/JNEUROSCI.2018-07.2007
Complement induction in spinal cord microglia results in anaphylatoxin C5a-mediated pain hypersensitivity
Abstract
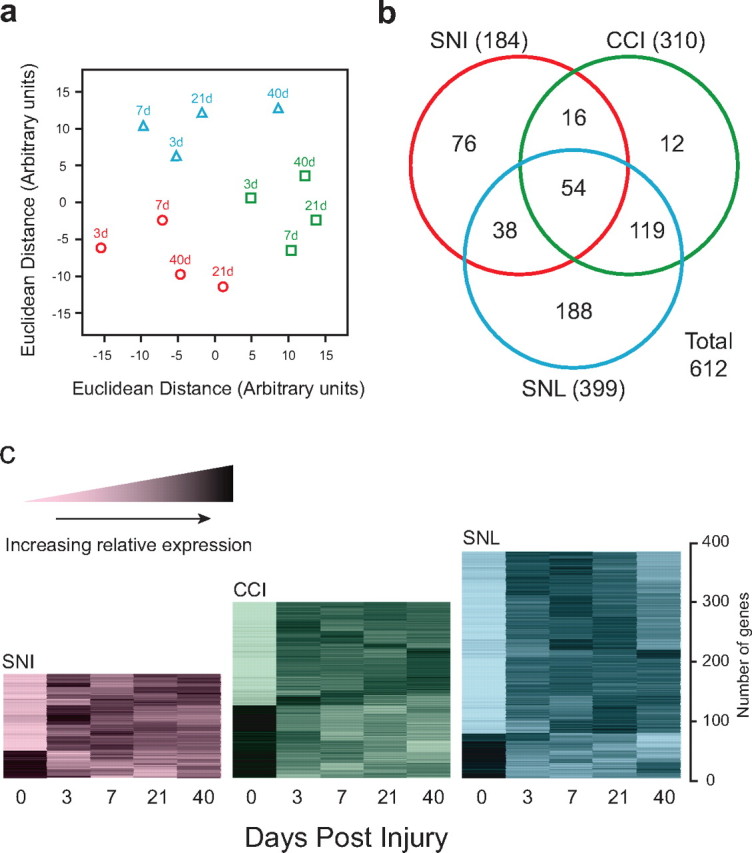
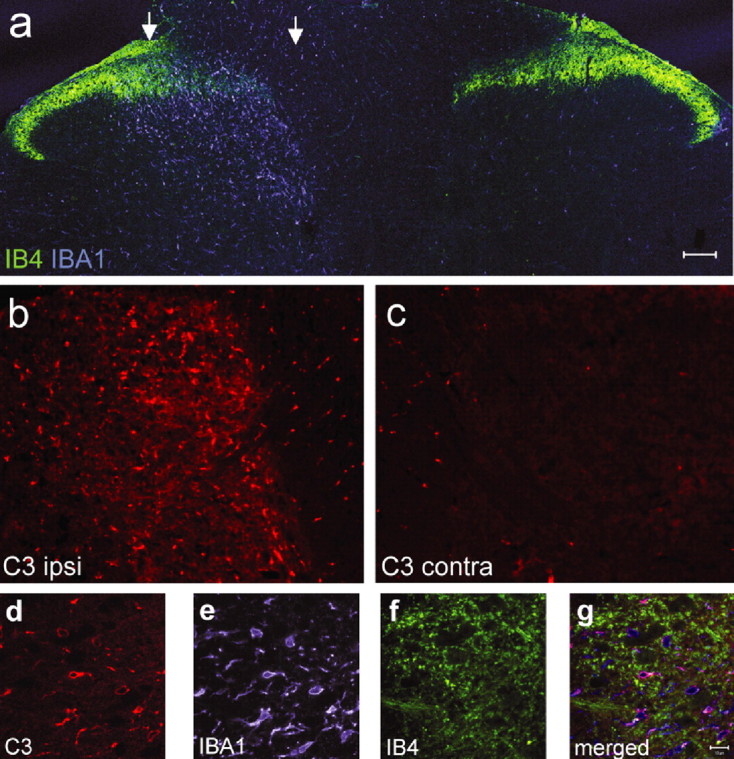
Microarray expression profiles reveal substantial changes in gene expression in the ipsilateral dorsal horn of the spinal cord in response to three peripheral nerve injury models of neuropathic pain. However, only 54 of the 612 regulated genes are commonly expressed across all the neuropathic pain models. Many of the commonly regulated transcripts are immune related and include the complement components C1q, C3, and C4, which we find are expressed only by microglia. C1q and C4 are, moreover, the most strongly regulated of all 612 regulated genes. In addition, we find that the terminal complement component C5 and the C5a receptor (C5aR) are upregulated in spinal microglia after peripheral nerve injury. Mice null for C5 had reduced neuropathic pain sensitivity, excluding C3a as a pain effector. C6-deficient rats, which cannot form the membrane attack complex, have a normal neuropathic pain phenotype. However, C5a applied intrathecally produces a dose-dependent, slow-onset cold pain in naive animals. Furthermore, a C5aR peptide antagonist reduces cold allodynia in neuropathic pain models. We conclude that induction of the complement cascade in spinal cord microglia after peripheral nerve injury contributes to neuropathic pain through the release and action of the C5a anaphylatoxin peptide.
Figures






References
-
- Arruda JL, Colburn RW, Rickman AJ, Rutkowski MD, DeLeo JA. Increase of interleukin-6 mRNA in the spinal cord following peripheral nerve injury in the rat: potential role of IL-6 in neuropathic pain. Brain Res Mol Brain Res. 1998;62:228–235. - PubMed
-
- Bennett GJ, Xie YK. A peripheral mononeuropathy in rat that produces disorders of pain sensation like those seen in man. Pain. 1988;33:87–107. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Molecular Biology Databases
Miscellaneous