

Identification of a responsible promoter region and a key transcription factor, CCAAT/enhancer-binding protein epsilon, for up-regulation of PHGPx in HL60 cells stimulated with TNF alpha
- PMID: 17688422
- PMCID: PMC2267347
- DOI: 10.1042/BJ20070245
Identification of a responsible promoter region and a key transcription factor, CCAAT/enhancer-binding protein epsilon, for up-regulation of PHGPx in HL60 cells stimulated with TNF alpha
Abstract
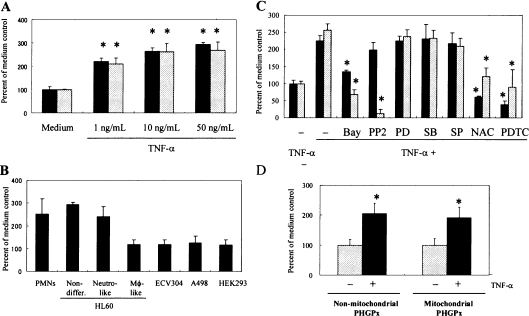
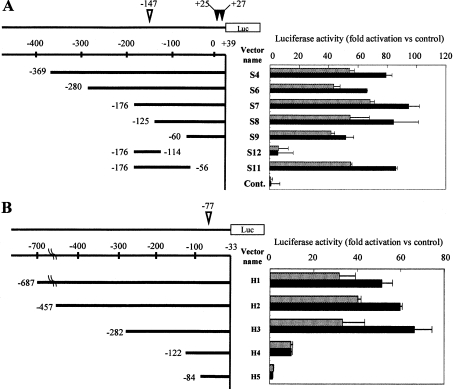
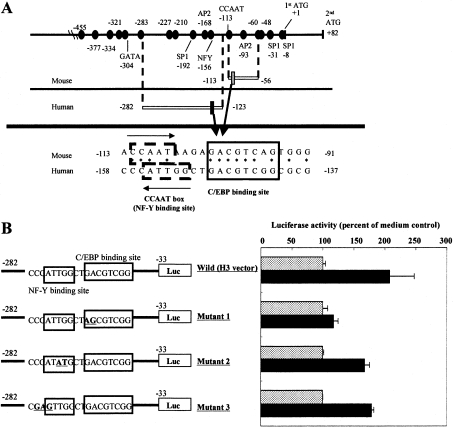
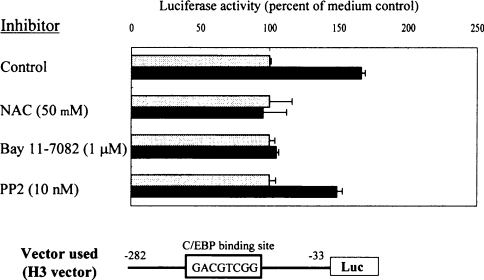
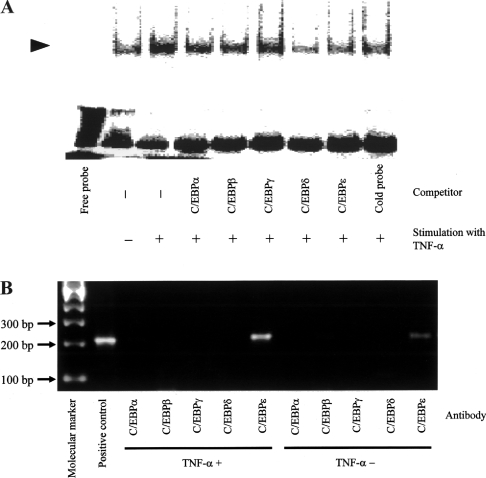
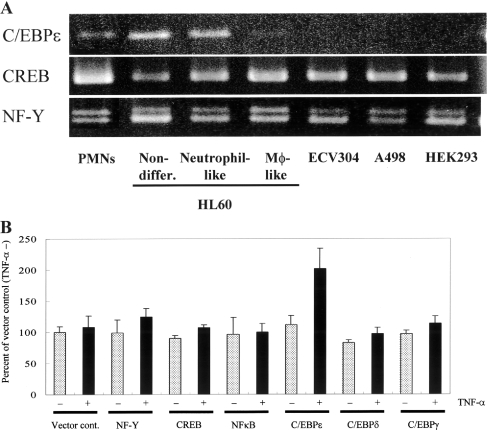
In the present study we investigated promoter regions of the PHGPx [phospholipid hydroperoxide GPx (glutathione peroxidase)] gene and transcription factors involved in TNFalpha (tumour necrosis factor alpha)-induced up-regulation of PHGPx in non-differentiated HL60 cells. Non-differentiated HL60 cells displayed up-regulation of non-mitochondrial and mitochondrial PHGPx mRNA in response to TNFalpha stimulation. The promoter activity was up-regulated by TNFalpha stimulation in cells transfected with a luciferase reporter vector encoding the region from -282 to -123 of the human PHGPx gene compared with the non-stimulated control. The up-regulated promoter activity was effectively abrogated by a mutation in the C/EBP (CCAAT/enhancer-binding protein)-binding sequence in this region. ChIP (chromatin immunoprecipitation) assays demonstrated that C/EBPepsilon bound to the -247 to -34 region in HL60 cells, but C/EBPalpha, beta, gamma and delta did not. The binding of C/EBPepsilon to the promoter region was increased in HL60 cells stimulated with TNFalpha compared with that of the non-stimulated control. An increased binding of nuclear protein to the C/EBP-binding sequence was observed by EMSA (electrophoretic mobility-shift assay) in cells stimulated with TNFalpha, and it was inhibited by pre-treatment with an anti-C/EBPepsilon antibody, but not with other antibodies. The C/EBPepsilon mRNA was expressed in PMNs (polymorphonuclear cells), non-differentiated HL60 cells and neutrophil-like differentiated HL60 cells displaying TNFalpha-induced up-regulation of PHGPx mRNA, but not in macrophage-like differentiated HL60 cells, HEK-293 cells (human embryonic kidney-293 cells) and other cell lines exhibiting no up-regulation. The up-regulation of PHGPx mRNA, however, was detected in HEK-293 cells overexpressing C/EBPepsilon as a result of TNFalpha stimulation. These results indicate that C/EBPepsilon is a critical transcription factor in TNFalpha-induced up-regulation of PHGPx expression.
Figures
References
-
- Root R. K., Cohen M. S. The microbicidal mechanisms of human neutrophils and eosinophils. Rev. Infect. Dis. 1981;3:565–598. - PubMed
-
- Babior B. M. Oxygen-dependent microbial killing by phagocytosis. N. Engl. J. Med. 1978;298:659–668. - PubMed
-
- Scapini P., Lapinet-Vera J. A., Gasperini S., Calzetti F., Bazzoni F., Cassatella M. A. The neutrophil as a cellular source of chemokines. Immunol. Rev. 2000;177:195–203. - PubMed
-
- Ford-Hutchinson A. W., Bray M. A., Doig M. V., Shipley M. E., Smith M. J. H. Leukotriene B, a potent chemokinetic and aggregating substance released from polymorphonuclear leukocytes. Nature. 1980;286:264–265. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Miscellaneous