

Early axonopathy preceding neurofibrillary tangles in mutant tau transgenic mice
- PMID: 17690183
- PMCID: PMC1959508
- DOI: 10.2353/ajpath.2007.070345
Early axonopathy preceding neurofibrillary tangles in mutant tau transgenic mice
Abstract
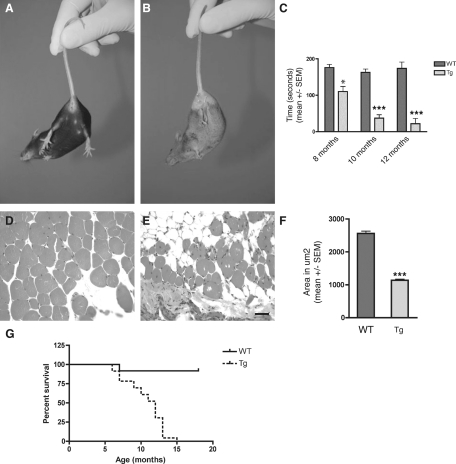
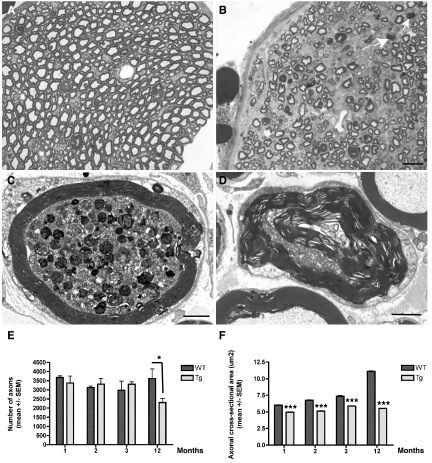
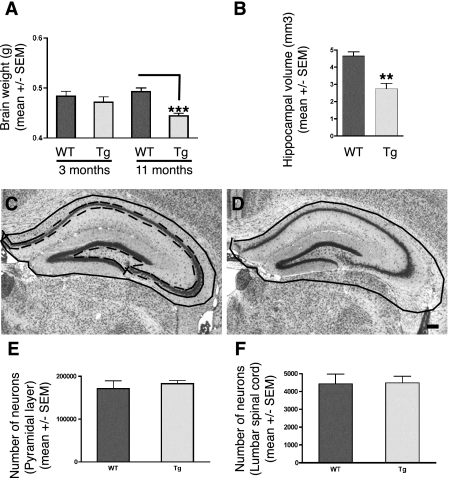
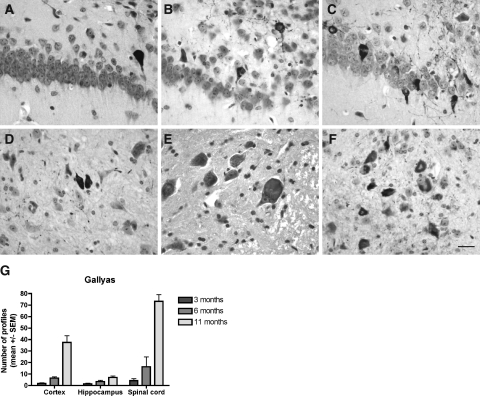
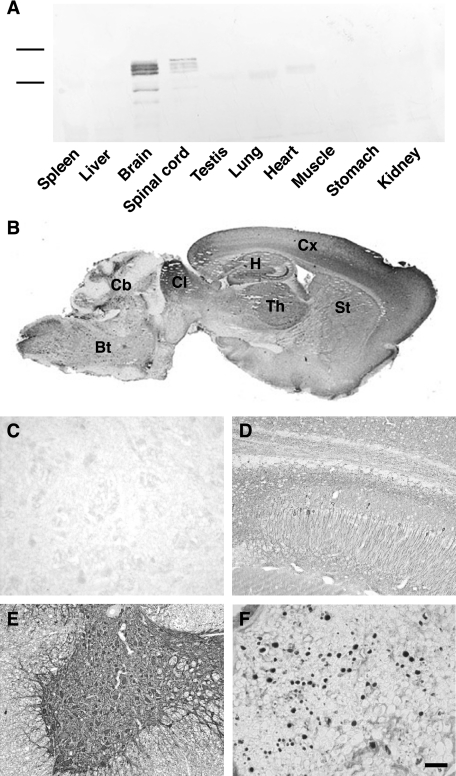
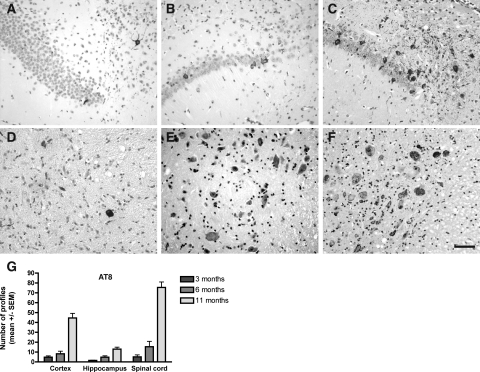
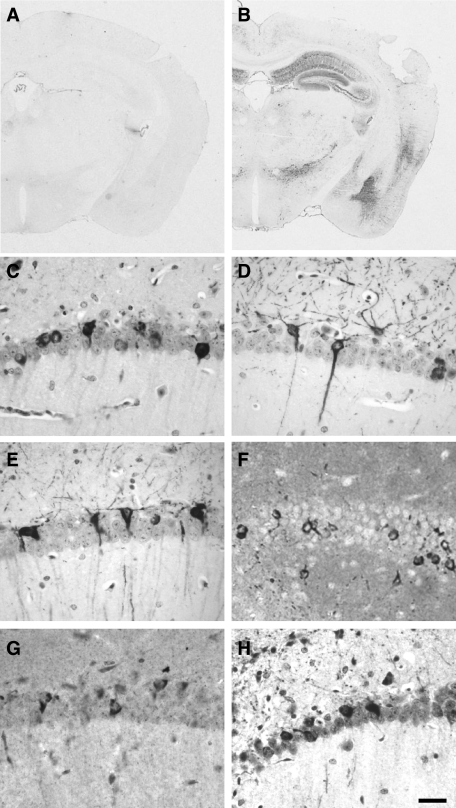
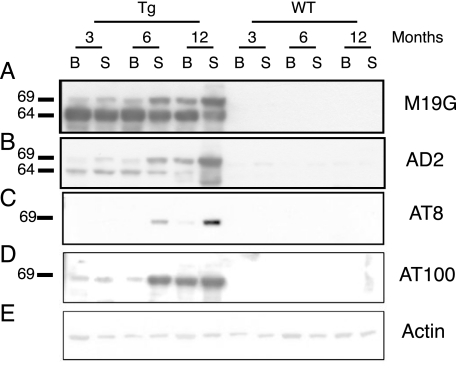
Neurodegenerative diseases characterized by brain and spinal cord involvement often show widespread accumulations of tau aggregates. We have generated a transgenic mouse line (Tg30tau) expressing in the forebrain and the spinal cord a human tau protein bearing two pathogenic mutations (P301S and G272V). These mice developed age-dependent brain and hippocampal atrophy, central and peripheral axonopathy, progressive motor impairment with neurogenic muscle atrophy, and neurofibrillary tangles and had decreased survival. Axonal spheroids and axonal atrophy developed early before neurofibrillary tangles. Neurofibrillary inclusions developed in neurons at 3 months and were of two types, suggestive of a selective vulnerability of neurons to form different types of fibrillary aggregates. A first type of tau-positive neurofibrillary tangles, more abundant in the forebrain, were composed of ribbon-like 19-nm-wide filaments and twisted paired helical filaments. A second type of tau and neurofilament-positive neurofibrillary tangles, more abundant in the spinal cord and the brainstem, were composed of 10-nm-wide neurofilaments and straight 19-nm filaments. Unbiased stereological analysis indicated that total number of pyramidal neurons and density of neurons in the lumbar spinal cord were not reduced up to 12 months in Tg30tau mice. This Tg30tau model thus provides evidence that axonopathy precedes tangle formation and that both lesions can be dissociated from overt neuronal loss in selected brain areas but not from neuronal dysfunction.
Figures
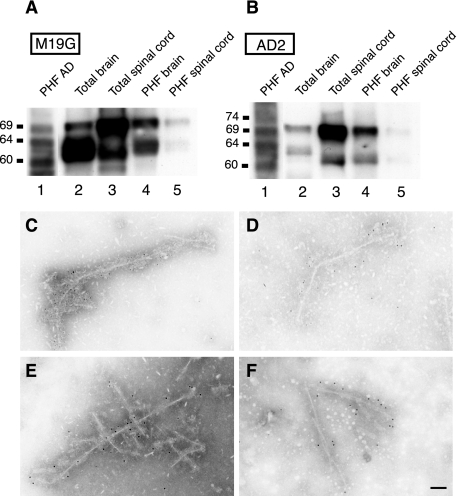
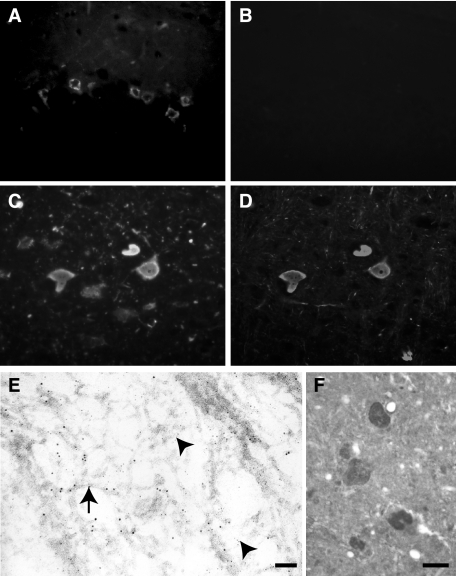
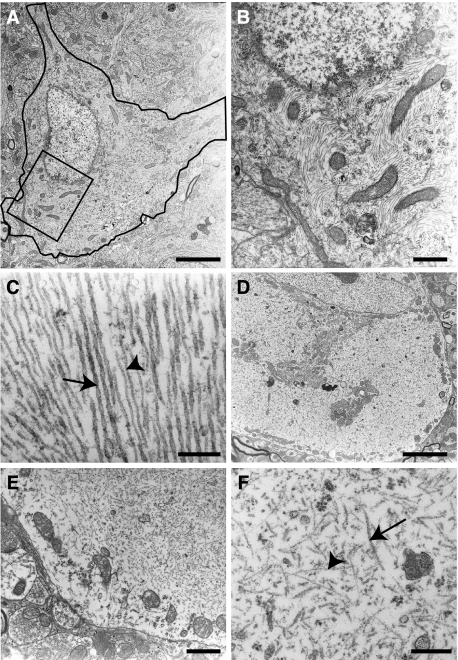
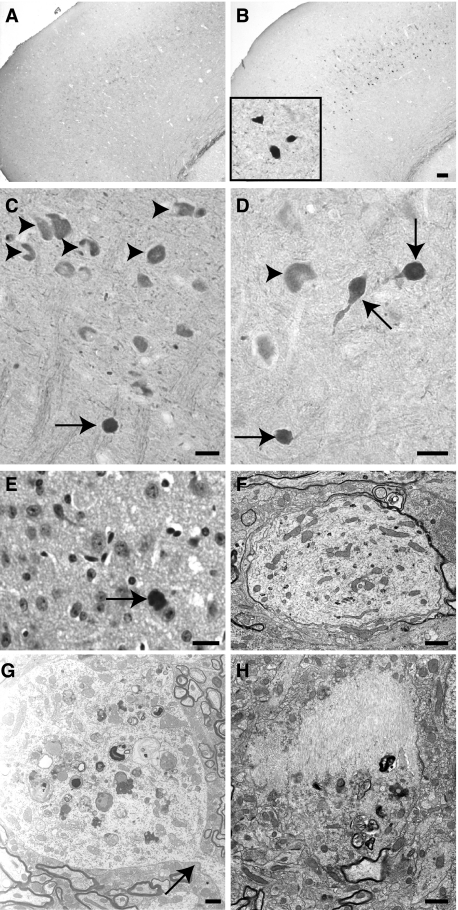
References
-
- Lee VMY, Goedert M, Trojanowski JQ. Neurodegenerative tauopathies. Annu Rev Neurosci. 2001;24:1121–1159. - PubMed
-
- Rodgers-Johnson P, Garruto RM, Yanagihara R, Chen KM, Gajdusek DC, Gibbs CJ., Jr Amyotrophic lateral sclerosis and parkinsonism-dementia on Guam: a 30-year evaluation of clinical and neuropathologic trends. Neurology. 1986;36:7–13. - PubMed
-
- Lomen-Hoerth C, Anderson T, Miller B. The overlap of amyotrophic lateral sclerosis and frontotemporal dementia. Neurology. 2002;59:1077–1079. - PubMed
-
- Buée L, Bussière T, Buée-Scherrer V, Delacourte A, Hof PR. Tau protein isoforms, phosphorylation and role in neurodegenerative disorders. Brain Res Rev. 2000;33:95–130. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
