

Impaired GABAergic transmission and altered hippocampal synaptic plasticity in collybistin-deficient mice
- PMID: 17690689
- PMCID: PMC1994120
- DOI: 10.1038/sj.emboj.7601819
Impaired GABAergic transmission and altered hippocampal synaptic plasticity in collybistin-deficient mice
Abstract
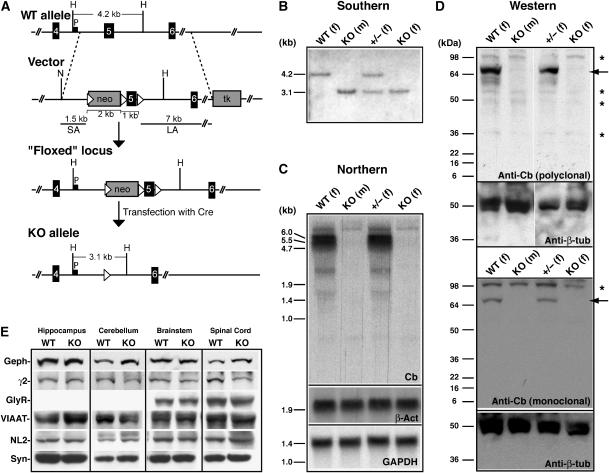
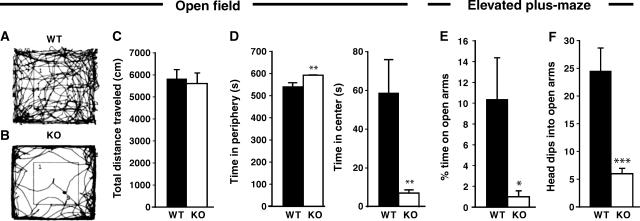
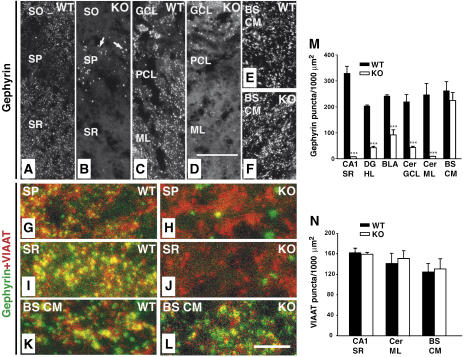
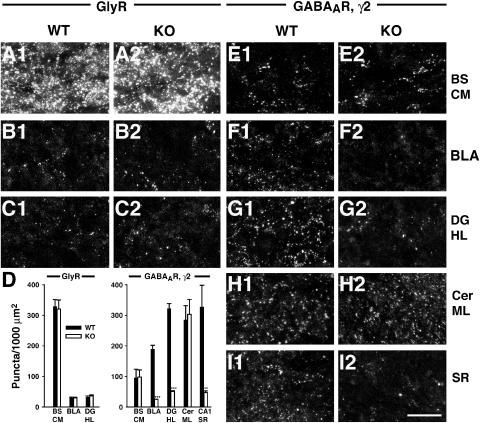
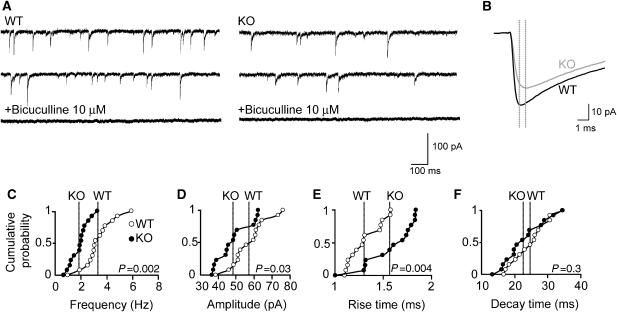
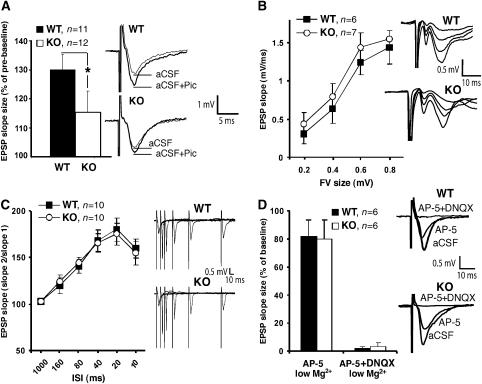
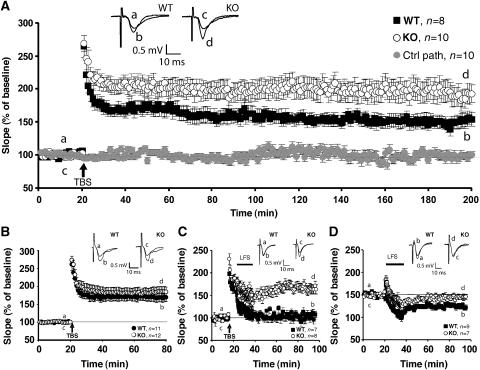
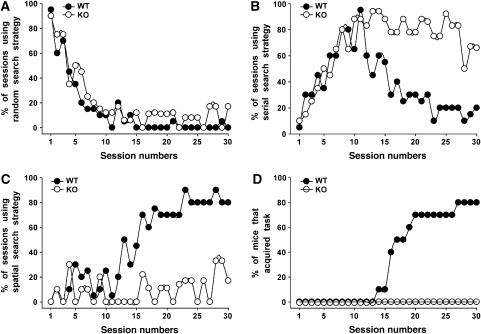
Collybistin (Cb) is a brain-specific guanine nucleotide exchange factor that has been implicated in plasma membrane targeting of the postsynaptic scaffolding protein gephyrin found at glycinergic and GABAergic synapses. Here we show that Cb-deficient mice display a region-specific loss of postsynaptic gephyrin and GABA(A) receptor clusters in the hippocampus and the basolateral amygdala. Cb deficiency is accompanied by significant changes in hippocampal synaptic plasticity, due to reduced dendritic GABAergic inhibition. Long-term potentiation is enhanced, and long-term depression reduced, in Cb-deficient hippocampal slices. Consistent with the anatomical and electrophysiological findings, the animals show increased levels of anxiety and impaired spatial learning. Together, our data indicate that Cb is essential for gephyrin-dependent clustering of a specific set of GABA(A) receptors, but not required for glycine receptor postsynaptic localization.
Figures
References
-
- Bach ME, Hawkins RD, Osman M, Kandel ER, Mayford M (1995) Impairment of spatial but not contextual memory in CaMKII mutant mice with a selective loss of hippocampal LTP in the range of the theta frequency. Cell 81: 905–915 - PubMed
-
- Barnes CA (1979) Memory deficits associated with senescence: a neurophysiological and behavioral study in the rat. J Comp Physiol Psychol 93: 74–104 - PubMed
-
- Bausen M, Fuhrmann JC, Betz H, O'Sullivan GA (2006) The state of the actin cytoskeleton determines its association with gephyrin: Role of ena/VASP family members. Mol Cell Neurosci 31: 376–386 - PubMed
-
- Bear MF, Abraham WC (1996) Long-term depression in hippocampus. Annu Rev Neurosci 19: 437–462 - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
