

Complexin I is required for mammalian sperm acrosomal exocytosis
- PMID: 17692307
- PMCID: PMC2099451
- DOI: 10.1016/j.ydbio.2007.07.009
Complexin I is required for mammalian sperm acrosomal exocytosis
Abstract
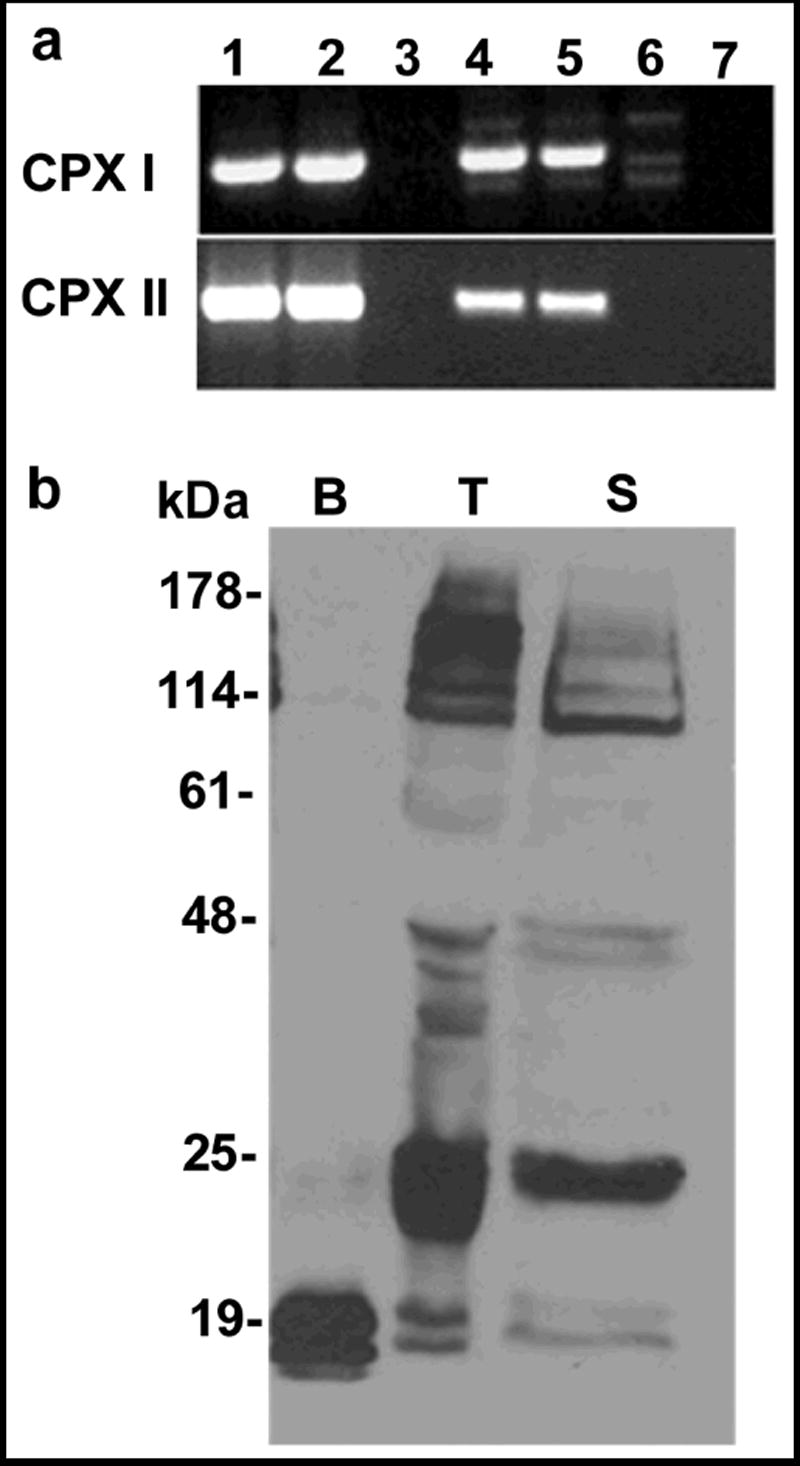
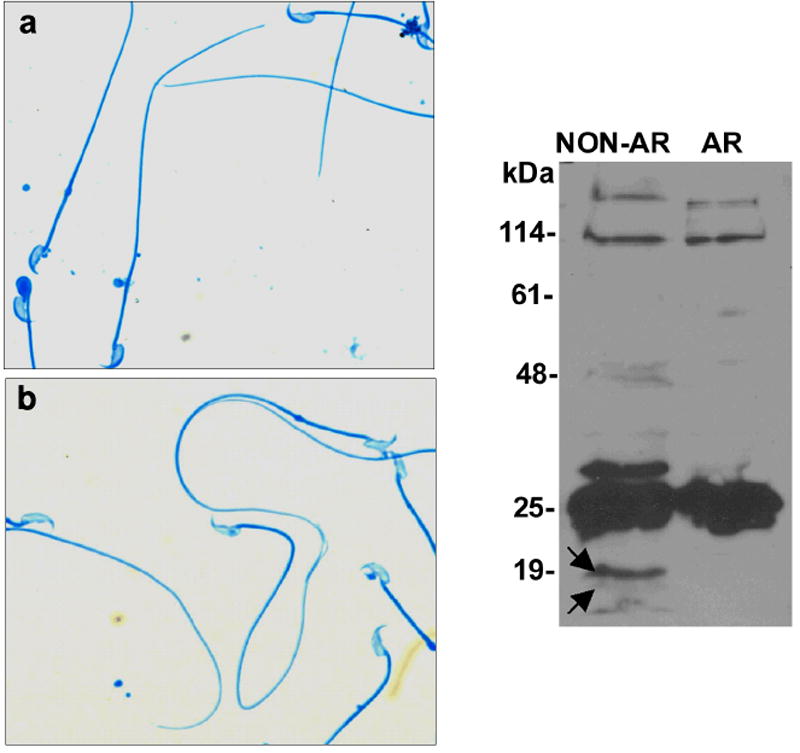
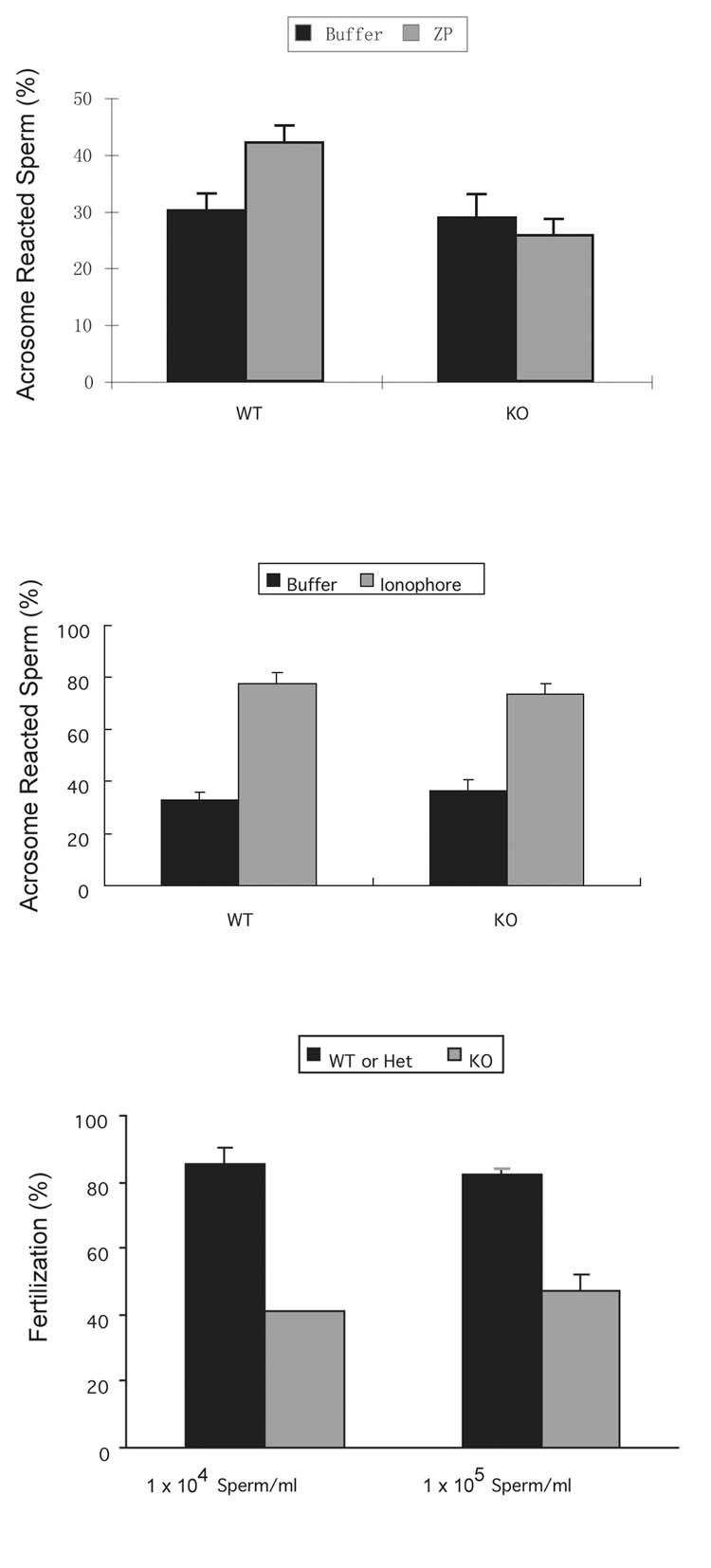
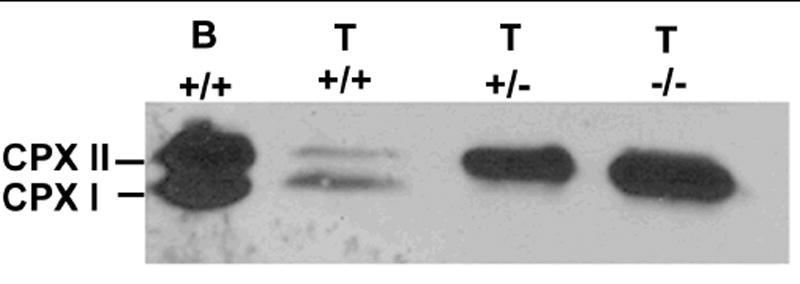
Regulated exocytosis in many cells is controlled by the SNARE complex, whose core includes three proteins that promote membrane fusion. Complexins I and II are highly related cytosolic proteins that bind tightly to the assembled SNARE complex and regulate neuronal exocytosis. Like somatic cells, sperm undergo regulated exocytosis; however, sperm release a single large vesicle, the acrosome, whose release has different characteristics than neuronal exocytosis. Acrosomal release is triggered upon sperm adhesion to the mammalian egg extracellular matrix (zona pellucida) to allow penetration of the egg coat. Membrane fusion occurs at multiple points within the acrosome but how fusion is activated and the formation and progression of fusion points is synchronized is unclear. We show that complexins I and II are found in acrosome-intact mature sperm, bind to SNARE complex proteins, and are not detected in sperm after acrosomal exocytosis (acrosome reaction). Although complexin-I-deficient sperm acrosome-react in response to calcium ionophore, they do not acrosome-react in response to egg zona pellucida proteins and have reduced fertilizing ability, in vitro. Complexin II is present in the complexin-I-deficient sperm and its expression is increased in complexin-I-deficient testes. Therefore, complexin I functions in exocytosis in two related but morphologically distinct secretory processes. Sperm are unusual because they express both complexins I and II but have a unique and specific requirement for complexin I.
Figures
Similar articles
-
Complexin-I-deficient sperm are subfertile due to a defect in zona pellucida penetration.Reproduction. 2008 Sep;136(3):323-34. doi: 10.1530/REP-07-0569. Epub 2008 Jun 24. Reproduction. 2008. PMID: 18577553
-
Involvement of complexin 2 in docking, locking and unlocking of different SNARE complexes during sperm capacitation and induced acrosomal exocytosis.PLoS One. 2012;7(3):e32603. doi: 10.1371/journal.pone.0032603. Epub 2012 Mar 6. PLoS One. 2012. PMID: 22412896 Free PMC article.
-
Dynamic resolution of acrosomal exocytosis in human sperm.J Cell Sci. 2008 Jul 1;121(Pt 13):2130-5. doi: 10.1242/jcs.030379. Epub 2008 Jun 3. J Cell Sci. 2008. PMID: 18522990
-
The role of the acrosomal matrix in fertilization.Int J Dev Biol. 2008;52(5-6):511-22. doi: 10.1387/ijdb.072532mb. Int J Dev Biol. 2008. PMID: 18649264 Review.
-
Mammalian sperm molecules that are potentially important in interaction with female genital tract and egg vestments.Zygote. 2001 Feb;9(1):51-69. doi: 10.1017/s096719940100106x. Zygote. 2001. PMID: 11273033 Review.
Cited by
-
Fertilization: a sperm's journey to and interaction with the oocyte.J Clin Invest. 2010 Apr;120(4):984-94. doi: 10.1172/JCI41585. Epub 2010 Apr 1. J Clin Invest. 2010. PMID: 20364096 Free PMC article. Review.
-
A multi-faceted approach to understanding male infertility: gene mutations, molecular defects and assisted reproductive techniques (ART).J Assist Reprod Genet. 2014 Sep;31(9):1115-37. doi: 10.1007/s10815-014-0280-6. Epub 2014 Aug 13. J Assist Reprod Genet. 2014. PMID: 25117645 Free PMC article. Review.
-
Intrinsically disordered proteins in synaptic vesicle trafficking and release.J Biol Chem. 2019 Mar 8;294(10):3325-3342. doi: 10.1074/jbc.REV118.006493. Epub 2019 Jan 30. J Biol Chem. 2019. PMID: 30700558 Free PMC article. Review.
-
Deep evolutionary origins of neurobiology: Turning the essence of 'neural' upside-down.Commun Integr Biol. 2009;2(1):60-5. doi: 10.4161/cib.2.1.7620. Commun Integr Biol. 2009. PMID: 19513267 Free PMC article.
-
PKA-dependent phosphorylation of LIMK1 and Cofilin is essential for mouse sperm acrosomal exocytosis.Dev Biol. 2015 Sep 15;405(2):237-49. doi: 10.1016/j.ydbio.2015.07.008. Epub 2015 Jul 10. Dev Biol. 2015. PMID: 26169470 Free PMC article.
References
-
- Bennett MK, Calakos N, Scheller RH. Syntaxin: a synaptic protein implicated in docking of synaptic vesicles at presynaptic active zones. Science. 1992;257:255–259. - PubMed
-
- Chen X, Tomchick DR, Kovrigin E, Arac D, Machius M, Sudhof TC, Rizo J. Three-dimensional structure of the complexin/SNARE complex. Neuron. 2002;33:397–409. - PubMed
-
- Choi YH, Toyoda Y. Cyclodextrin removes cholesterol from mouse sperm and induces capacitation in a protein-free medium. Biol Reprod. 1998;59:1328–33. - PubMed
-
- Fasshauer D, Eliason WK, Brunger AT, Jahn R. Identification of a minimal core of the synaptic SNARE complex sufficient for reversible assembly and disassembly. Biochemistry. 1998;37:10354–62. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
