

Fluctuations of transfer RNAs between classical and hybrid states
- PMID: 17693476
- PMCID: PMC2072051
- DOI: 10.1529/biophysj.107.109884
Fluctuations of transfer RNAs between classical and hybrid states
Abstract
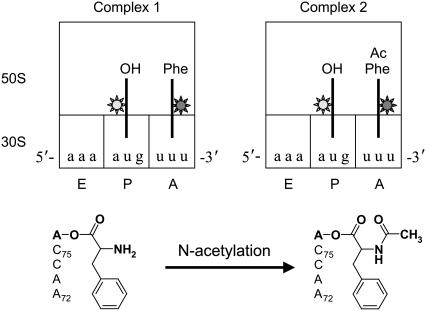
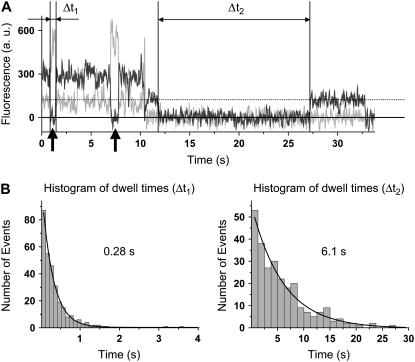
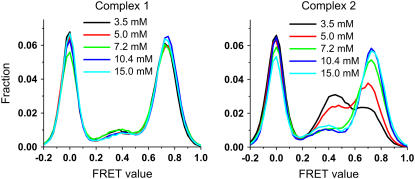
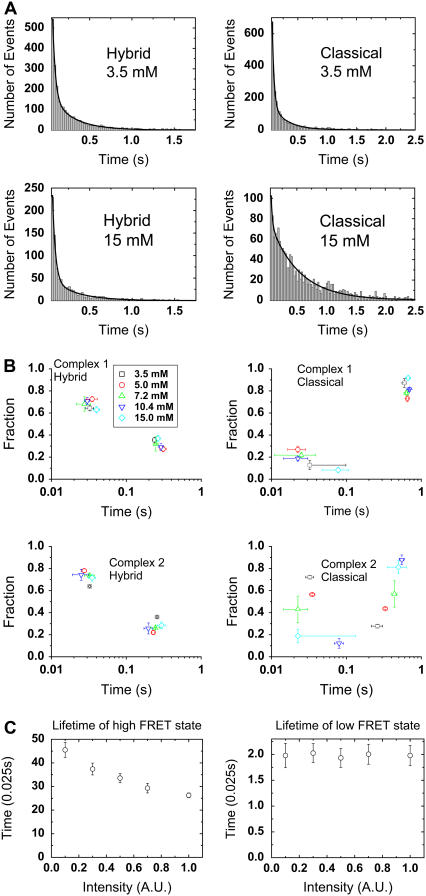
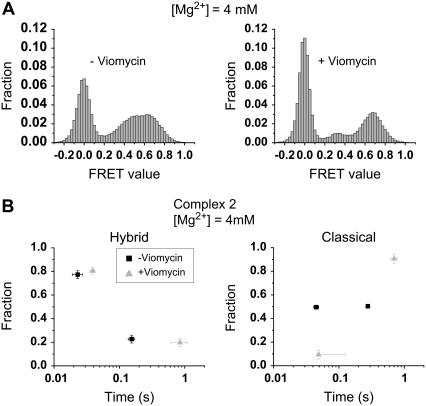
Adjacent transfer RNAs (tRNAs) in the A- and P-sites of the ribosome are in dynamic equilibrium between two different conformations called classical and hybrid states before translocation. Here, we have used single-molecule fluorescence resonance energy transfer to study the effect of Mg(2+) on tRNA dynamics with and without an acetyl group on the A-site tRNA. When the A-site tRNA is not acetylated, tRNA dynamics do not depend on [Mg(2+)], indicating that the relative positions of the substrates for peptide-bond formation are not affected by Mg(2+). In sharp contrast, when the A-site tRNA is acetylated, Mg(2+) lengthens the lifetime of the classical state but does not change the lifetime of the hybrid state. Based on these findings, the classical state resembles a state with direct stabilization of tertiary structure by Mg(2+) ions whereas the hybrid state resembles a state with little Mg(2+)-assisted stabilization. The antibiotic viomycin, a translocation inhibitor, suppresses tRNA dynamics, suggesting that the enhanced fluctuations of tRNAs after peptide-bond formation drive spontaneous attempts at translocation by the ribosome.
Figures
References
-
- Semenkov, Y. P., M. V. Rodnina, and W. Wintermeyer. 2000. Energetic contribution of tRNA hybrid state formation to translocation catalysis on the ribosome. Nat. Struct. Biol. 7:1027–1031. - PubMed
-
- Studer, S. M., J. S. Feinberg, and S. Joseph. 2003. Rapid kinetic analysis of EF-G-dependent mRNA translocation in the ribosome. J. Mol. Biol. 327:369–381. - PubMed
-
- Fredrick, K., and H. F. Noller. 2002. Accurate translocation of mRNA by the ribosome requires a peptidyl group or its analog on the tRNA moving into the 30S P site. Mol. Cell. 9:1125–1131. - PubMed
-
- Moazed, D., and H. F. Noller. 1989. Interaction of transfer-RNA with 23s ribosomal-RNA in the ribosomal A-sites, P-sites, and E-sites. Cell. 57:585–597. - PubMed
-
- Moazed, D., and H. F. Noller. 1989. Intermediate states in the movement of transfer-RNA in the ribosome. Nature. 342:142–148. - PubMed
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
