

Consequences of parameter differences in a model of short-term persistent spiking buffers provided by pyramidal cells in entorhinal cortex
- PMID: 17698043
- PMCID: PMC2722951
- DOI: 10.1016/j.brainres.2007.06.067
Consequences of parameter differences in a model of short-term persistent spiking buffers provided by pyramidal cells in entorhinal cortex
Abstract
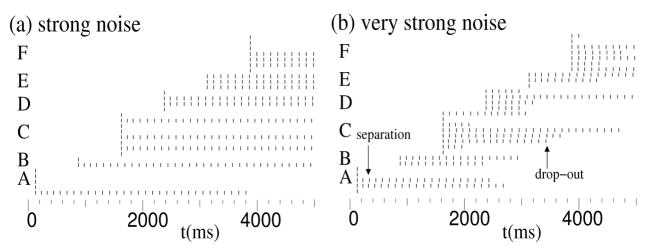
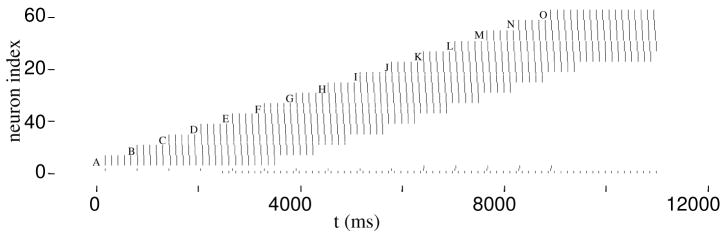
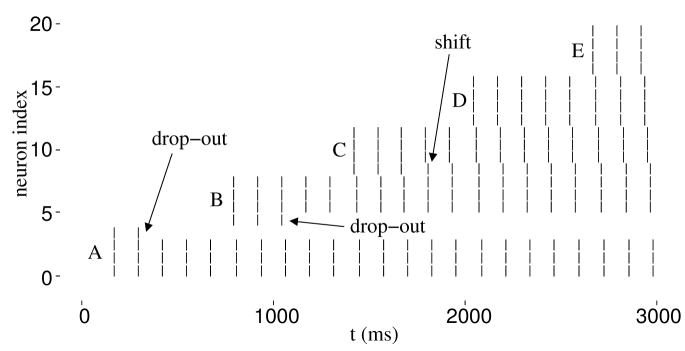
In previous simulations of hippocampus-dependent and prefrontal cortex-dependent tasks, we demonstrated the use of one-shot short-term buffering with time compression that may be achieved through persistent spiking activity during theta rhythm. A biophysically plausible implementation of such a first-in first-out buffer of short sequences of spike patterns includes noise and differences between the parameter values of individual model pyramidal cells. We show that a specific set of parameters determines model buffer capacity and buffer function, and individual differences can have consequences similar to those of noise. The set of parameters includes the frequency of network theta rhythm and the strength of recurrent inhibition (affecting capacity), as well as the time constants of the characteristic after-depolarizing response and the phase of afferent input during theta rhythm (affecting buffer function). Given a sufficient number of pyramidal cells in layer II of entorhinal cortex, and in each self-selected category of pyramidal cells with similar model parameters, buffer function within a category is reliable with category-specific properties. Properties include buffering of spikes in the order of inputs or in the reversed order. Multiple property sets may enable parallel buffers with different capacities, which may underlie differences of place field sizes and may interact with grid cell firing in a separate population of layer II stellate cells in the entorhinal cortex.
Figures









Similar articles
-
First-in-first-out item replacement in a model of short-term memory based on persistent spiking.Cereb Cortex. 2007 Aug;17(8):1766-81. doi: 10.1093/cercor/bhl088. Epub 2006 Oct 9. Cereb Cortex. 2007. PMID: 17030561
-
A computational study on plasticity during theta cycles at Schaffer collateral synapses on CA1 pyramidal cells in the hippocampus.Hippocampus. 2015 Feb;25(2):208-18. doi: 10.1002/hipo.22365. Epub 2014 Sep 25. Hippocampus. 2015. PMID: 25220633
-
Grid cell mechanisms and function: contributions of entorhinal persistent spiking and phase resetting.Hippocampus. 2008;18(12):1213-29. doi: 10.1002/hipo.20512. Hippocampus. 2008. PMID: 19021258 Free PMC article.
-
Linking cellular mechanisms to behavior: entorhinal persistent spiking and membrane potential oscillations may underlie path integration, grid cell firing, and episodic memory.Neural Plast. 2008;2008:658323. doi: 10.1155/2008/658323. Neural Plast. 2008. PMID: 18670635 Free PMC article. Review.
-
Dual phase and rate coding in hippocampal place cells: theoretical significance and relationship to entorhinal grid cells.Hippocampus. 2005;15(7):853-66. doi: 10.1002/hipo.20115. Hippocampus. 2005. PMID: 16145693 Free PMC article. Review.
Cited by
-
A working memory model for serial order that stores information in the intrinsic excitability properties of neurons.J Comput Neurosci. 2013 Oct;35(2):187-99. doi: 10.1007/s10827-013-0447-7. Epub 2013 Mar 29. J Comput Neurosci. 2013. PMID: 23539088
-
The cognitive neuroscience of working memory: relevance to CNTRICS and schizophrenia.Biol Psychiatry. 2008 Jul 1;64(1):11-7. doi: 10.1016/j.biopsych.2008.03.003. Epub 2008 Apr 8. Biol Psychiatry. 2008. PMID: 18400207 Free PMC article.
-
Reversed and forward buffering of behavioral spike sequences enables retrospective and prospective retrieval in hippocampal regions CA3 and CA1.Neural Netw. 2008 Mar-Apr;21(2-3):276-88. doi: 10.1016/j.neunet.2007.12.029. Epub 2007 Dec 28. Neural Netw. 2008. PMID: 18242057 Free PMC article.
-
A robust in vivo-like persistent firing supported by a hybrid of intracellular and synaptic mechanisms.PLoS One. 2015 Apr 22;10(4):e0123799. doi: 10.1371/journal.pone.0123799. eCollection 2015. PLoS One. 2015. PMID: 25901969 Free PMC article.
References
-
- Acker C, Kopell N, White J. Synchronization of strongly coupled excitatory neurons: relating network behavior to biophysics. Journal of Computational Neuroscience. 2003;15(1):71–90. - PubMed
-
- Andrade R. The effect of carbachol which affects muscarinic receptors was investigated in prefrontal layer v neurons. Brain Research. 1991;541:81–93. - PubMed
-
- Atkinson R, Shiffrin R. Human memory: A proposed system and its control processes. In: Spence K, Spence J, editors. The psychology of learning and motivation. Vol. 2. New York: Academic Press.; 1968. pp. 89–105.
-
- Cannon R, Hasselmo M, Koene R. From biophysics to behaviour: Cata-comb2 and the design of biologically plausable models for spatial navigation. Neuroinformatics. 2003;1(1):3–42. - PubMed
Publication types
MeSH terms
Grants and funding
LinkOut - more resources
Full Text Sources
