

doi: 10.1073/pnas.0703078104.
Epub 2007 Aug 14.
Yeast aconitase binds and provides metabolically coupled protection to mitochondrial DNA
Affiliations
- PMID: 17698960
- PMCID: PMC1959452
- DOI: 10.1073/pnas.0703078104
Item in Clipboard
Yeast aconitase binds and provides metabolically coupled protection to mitochondrial DNA
Proc Natl Acad Sci U S A.
.
Abstract
Aconitase (Aco1p) is a multifunctional protein: It is an enzyme of the tricarboxylic acid cycle. In animal cells, Aco1p also is a cytosolic protein binding to mRNAs to regulate iron metabolism. In yeast, Aco1p was identified as a component of mtDNA nucleoids. Here we show that yeast Aco1p protects mtDNA from excessive accumulation of point mutations and ssDNA breaks and suppresses reductive recombination of mtDNA. Aconitase binds to both ds- and ssDNA, with a preference for GC-containing sequences. Therefore, mitochondria are opportunistic organelles that seize proteins, such as metabolic enzymes, for construction of the nucleoid, an mtDNA maintenance/segregation apparatus.
Conflict of interest statement
The authors declare no conflict of interest.
Figures
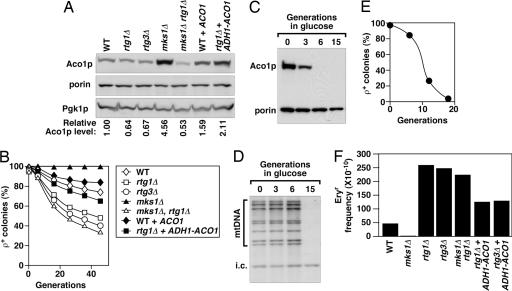
Aco1p levels are tightly correlated with the stability and mutation rate of ρ+ mtDNA. (A) Steady-state level of Aco1p. The level of porin is shown as a marker for mitochondrial proteins, and Aco1p levels relative to the wild-type control are normalized against the cytosolic Pgk1p. (B) Stability of the ρ+ genome in cells expressing different levels of Aco1p. mtDNA stability is expressed as a percentage of respiratory-competent (ρ+) colonies after growth in YPD for the number of generations as indicated. (C) Depletion of Aco1p in strain CS745–2C (aco1Δ::kan, ade2::GAL10-ACO1-URA3) during glucose repression in YPD medium. (D) Southern blot analysis of CfoI-digested total DNA probed for mtDNA in CS745–2C cells grown in YPD medium (i.c., internal control for sample loading). (E) Frequency of respiratory-competent cells in CS745–2C after growth in liquid YPD. The ρ+ cells were scored by the formation of red colonies when plated on YPGal medium. (F) Mutation rate in the ρ+ genome as expressed by the frequency of erythromycin-resistant (Eryr) colonies.
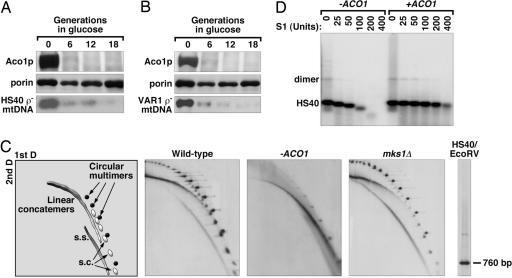
A role for Aco1p in the protection and maintenance of ρ− genomes. (A and B) Level of the HS40 and VAR1 ρ− mtDNA upon depletion of Aco1p in CS776/1 [aco1Δ, ade2::GAL10-ACO1-URA3, (HS40)] and CS777/2 [aco1Δ, ade2::GAL10-ACO1-URA3, (VAR1)] in glucose medium. Total DNA was digested with EcoRV and HincII and subjected to Southern blot by using probes specific for the HS40 and VAR1 ρ− genomes, respectively. Results of Western blot analysis are included to show the steady-state level of Aco1p and porin in the cells. (C) 2DAGE analysis of the HS40 ρ− mtDNA in cells depleted for Aco1p (−ACO1) or overexpressing ACO1 (mks1Δ), compared with cells expressing wild-type ACO1. (Left) Position of circular multimer, linear concatemer, supercoiled (s.c.), and the single-stranded (s.s.) forms of mtDNA. (Right) Position of the EcoRV-digested HS40 genome in the second-dimension electrophoresis. (D) S1 nuclease sensitivity of the HS40 ρ− genome extracted from cells with or without depletion of Aco1p.
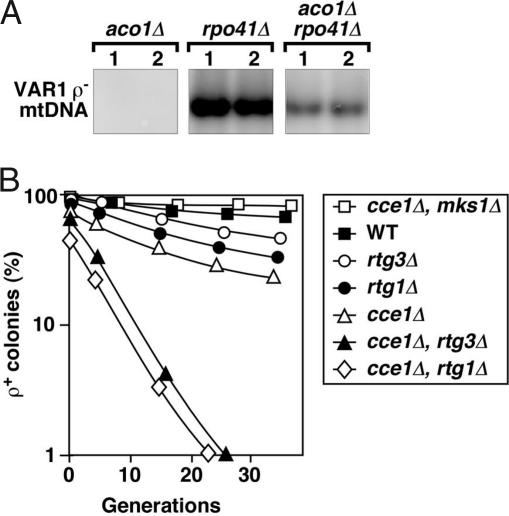
mtDNA stability in Aco1p-depleted cells disrupted for RPO41 and CCE1. (A) Southern blot analysis showing the level of the VAR1 ρ− mtDNA in representative cytoductants with aco1Δ or rpo41Δ or aco1Δ plus rpo41Δ. (B) Frequency of respiratory-competent cells in cce1Δ mutants in which the RTG pathway was up- or down-regulated.
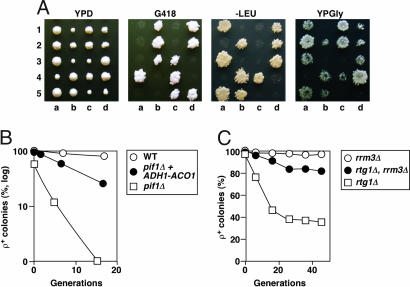
Epistasis between ACO1 expression and PIF1. (A) Growth phenotype of five complete tetrads from CS956 (pif1Δ::kan/+, mks1Δ::LEU2/+). (B) Frequency of respiratory-competent cells after growth in YPD medium. (C) Suppression of mtDNA instability in rtg1Δ mutant by rrm3Δ.
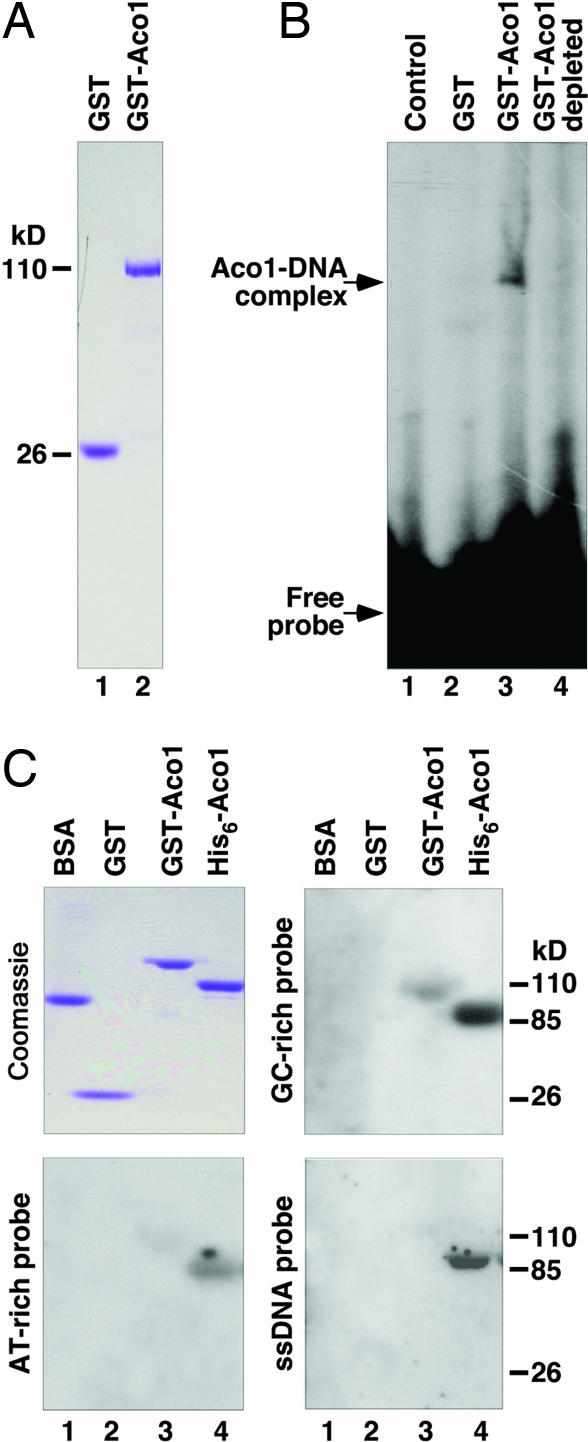
Aco1p binds to DNA. (A) Coomassie-stained gel showing purified GST and GST-Aco1p. (B) Gel retardation assay showing the binding of GST-Aco1 to the AT-rich dsDNA probe, HS40ATFR (SI Text ). Lane 1, no protein; lane 2, GST (2 μg); lane 3, GST-Aco1p (2 μg); lane 4, equal volume of aliquot after GST-Aco1p depletion by glutathione Sepharose 4B beads. (C) Coomassie-stained gel showing purified GST-Aco1p and His6-Aco1p, and Southwestern blot showing the binding of the fusion proteins to GC- and AT-rich ds- and ssDNA.
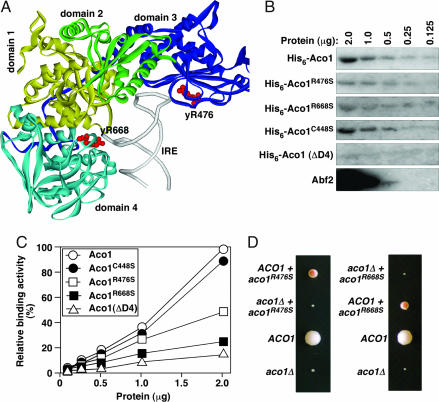
Aco1p variants compromised in DNA-binding in vitro are defective in mtDNA maintenance in vivo. (A) The position of Arg-541 and Arg-780 in rabbit IRP1 (equivalent to yArg476 and yArg668) in the IRE-binding conformation (3). (B) Southwestern blot showing the binding of Aco1p variants and Abf2 to the GC-rich BA box probe. Abf2p is loaded at a concentration 20-fold lower than the Aco1p variants. (C) Relative affinity of Aco1p variants to the GC-rich B and A box probe. (D) Representative tetrads showing that the meiotic segregants expressing only aco1R476S or aco1R668S form petite colonies on complete raffinose medium.
Similar articles
-
Aconitase couples metabolic regulation to mitochondrial DNA maintenance.Science. 2005 Feb 4;307(5710):714-7. doi: 10.1126/science.1106391. Science. 2005. PMID: 15692048
-
Mitochondrial DNA, aconitase 'wraps' it up.Trends Biochem Sci. 2005 Jun;30(6):294-6. doi: 10.1016/j.tibs.2005.04.007. Trends Biochem Sci. 2005. PMID: 15950872 Review.
-
Irc3 is a mitochondrial DNA branch migration enzyme.Sci Rep. 2016 May 19;6:26414. doi: 10.1038/srep26414. Sci Rep. 2016. PMID: 27194389 Free PMC article.
-
Mitochondrial DNA instability in cells lacking aconitase correlates with iron citrate toxicity.Oxid Med Cell Longev. 2013;2013:493536. doi: 10.1155/2013/493536. Epub 2013 Aug 26. Oxid Med Cell Longev. 2013. PMID: 24066190 Free PMC article.
-
Yeast and human mitochondrial helicases.Biochim Biophys Acta. 2013 Aug;1829(8):842-53. doi: 10.1016/j.bbagrm.2013.02.009. Epub 2013 Feb 27. Biochim Biophys Acta. 2013. PMID: 23454114 Review.
Cited by
-
Kinetic and Regulatory Properties of Yarrowia lipolytica Aconitate Hydratase as a Model-Indicator of Cell Redox State under pH Stress.Int J Mol Sci. 2023 Apr 21;24(8):7670. doi: 10.3390/ijms24087670. Int J Mol Sci. 2023. PMID: 37108831 Free PMC article.
-
Nitric Oxide Improves the Tolerance of Pleurotus ostreatus to Heat Stress by Inhibiting Mitochondrial Aconitase.Appl Environ Microbiol. 2020 Feb 18;86(5):e02303-19. doi: 10.1128/AEM.02303-19. Print 2020 Feb 18. Appl Environ Microbiol. 2020. PMID: 31862720 Free PMC article.
-
Aconitase regulation of erythropoiesis correlates with a novel licensing function in erythropoietin-induced ERK signaling.PLoS One. 2011;6(8):e23850. doi: 10.1371/journal.pone.0023850. Epub 2011 Aug 22. PLoS One. 2011. PMID: 21887333 Free PMC article.
-
Mutations in the dimer interface of dihydrolipoamide dehydrogenase promote site-specific oxidative damages in yeast and human cells.J Biol Chem. 2011 Nov 18;286(46):40232-45. doi: 10.1074/jbc.M111.274415. Epub 2011 Sep 19. J Biol Chem. 2011. PMID: 21930696 Free PMC article.
-
A quantitative screen for metabolic enzyme structures reveals patterns of assembly across the yeast metabolic network.Mol Biol Cell. 2019 Oct 1;30(21):2721-2736. doi: 10.1091/mbc.E19-04-0224. Epub 2019 Sep 4. Mol Biol Cell. 2019. PMID: 31483745 Free PMC article.
References
-
- Rouault TA. Nat Chem Biol. 2006;2:406–414. - PubMed
-
- Dupuy J, Volbeda A, Carpentier P, Darnault C, Moulis JM, Fontecilla-Camps JC. Structure (London) 2006;14:129–139. - PubMed
-
- Walden WE, Selezneva AI, Dupuy J, Volbeda A, Fontecilla-Camps JC, Theil EC, Volz K. Science. 2006;314:1903–1908. - PubMed
-
- Chen XJ, Wang X, Kaufman BA, Butow RA. Science. 2005;307:714–717. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Miscellaneous
