

Early antibodies specific for the neutralizing epitope on the receptor binding subunit of the lymphocytic choriomeningitis virus glycoprotein fail to neutralize the virus
- PMID: 17699567
- PMCID: PMC2168768
- DOI: 10.1128/JVI.00955-07
Early antibodies specific for the neutralizing epitope on the receptor binding subunit of the lymphocytic choriomeningitis virus glycoprotein fail to neutralize the virus
Abstract
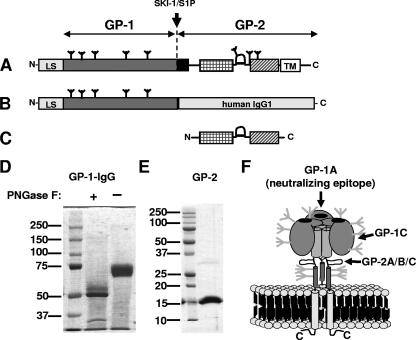
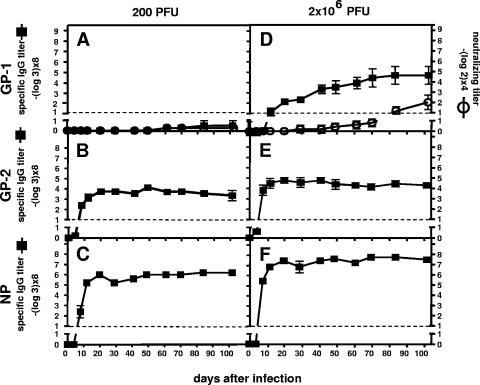
Lymphocytic choriomeningitis virus (LCMV) is a murine arenavirus whose glycoprotein consists of a transmembrane subunit (GP-2) and a receptor-binding subunit (GP-1). LCMV-neutralizing antibodies (nAbs) are directed against a single site on GP-1 and occur 1 month after the infection of cytotoxic-T-lymphocyte (CTL) deficient mice. In wild-type mice, however, CTLs control early infection, and weak nAb titers emerge very late (after 70 to 150 days) if at all. Production of recombinant GP-1 in native conformation enabled us to study the emergence of GP-1-binding antibodies directed against the neutralizing epitope. By combining binding and neutralization assays, we correlated the development of binding antibodies versus nAbs in wild-type and CTL-deficient mice after infection with different LCMV doses. We found that wild-type mice developed GP-1-specific antibodies already by day 8 after exposure to high but not low doses, demonstrating that naive GP-1-specific B cells were infrequent. Furthermore, the induced antibodies bound to the neutralizing GP-1 epitope but failed to neutralize the virus and therefore were of low affinity. In CTL-deficient mice, where massive viremia quickly levels initial differences in viral load, low and high doses induced low-affinity non-neutralizing GP-1-binding antibodies with kinetics similar to high-dose-infected wild-type mice. Only in CTL-deficient mice, however, the GP-1-specific antibodies developed into nAbs within 1 month. We conclude that LCMV uses a dual strategy to evade nAb responses in wild-type mice. First, LCMV exploits a "hole" in the murine B-cell repertoire, which provides only a small and narrow initial pool of low-affinity GP-1-specific B cells. Second, affinity maturation of the available low-affinity non-neutralizing antibodies is impaired.
Figures
Similar articles
-
Impairment and delay of neutralizing antiviral antibody responses by virus-specific cytotoxic T cells.J Immunol. 1993 Nov 15;151(10):5408-15. J Immunol. 1993. PMID: 7693811
-
Anti-viral protection and prevention of lymphocytic choriomeningitis or of the local footpad swelling reaction in mice by immunization with vaccinia-recombinant virus expressing LCMV-WE nucleoprotein or glycoprotein.Eur J Immunol. 1989 Mar;19(3):417-24. doi: 10.1002/eji.1830190302. Eur J Immunol. 1989. PMID: 2468501
-
Low-affinity cytotoxic T-lymphocytes require IFN-gamma to clear an acute viral infection.Virology. 1997 Mar 17;229(2):349-59. doi: 10.1006/viro.1997.8442. Virology. 1997. PMID: 9126248
-
Public, private and non-specific antibodies induced by non-cytopathic viral infections.Curr Opin Microbiol. 2004 Aug;7(4):426-33. doi: 10.1016/j.mib.2004.06.008. Curr Opin Microbiol. 2004. PMID: 15358263 Review.
-
Viral escape from the neutralizing antibody response: the lymphocytic choriomeningitis virus model.Immunogenetics. 2001 Apr;53(3):185-9. doi: 10.1007/s002510100314. Immunogenetics. 2001. PMID: 11398962 Review.
Cited by
-
Most neutralizing human monoclonal antibodies target novel epitopes requiring both Lassa virus glycoprotein subunits.Nat Commun. 2016 May 10;7:11544. doi: 10.1038/ncomms11544. Nat Commun. 2016. PMID: 27161536 Free PMC article.
-
"Negative vaccination" by specific CD4 T cell tolerisation enhances virus-specific protective antibody responses.PLoS One. 2007 Nov 14;2(11):e1162. doi: 10.1371/journal.pone.0001162. PLoS One. 2007. PMID: 18000535 Free PMC article.
-
Perturbation of Thymocyte Development Underlies the PRRS Pandemic: A Testable Hypothesis.Front Immunol. 2019 May 15;10:1077. doi: 10.3389/fimmu.2019.01077. eCollection 2019. Front Immunol. 2019. PMID: 31156633 Free PMC article.
-
Replication-incompetent rabies virus vector harboring glycoprotein gene of lymphocytic choriomeningitis virus (LCMV) protects mice from LCMV challenge.PLoS Negl Trop Dis. 2018 Apr 16;12(4):e0006398. doi: 10.1371/journal.pntd.0006398. eCollection 2018 Apr. PLoS Negl Trop Dis. 2018. PMID: 29659579 Free PMC article.
-
Xenoantigen-Dependent Complement-Mediated Neutralization of Lymphocytic Choriomeningitis Virus Glycoprotein-Pseudotyped Vesicular Stomatitis Virus in Human Serum.J Virol. 2019 Aug 28;93(18):e00567-19. doi: 10.1128/JVI.00567-19. Print 2019 Sep 15. J Virol. 2019. PMID: 31243134 Free PMC article.
References
-
- Althage, A., B. Odermatt, D. Moskophidis, T. Kundig, U. Hoffman-Rohrer, H. Hengartner, and R. M. Zinkernagel. 1992. Immunosuppression by lymphocytic choriomeningitis virus infection: competent effector T and B cells but impaired antigen presentation. Eur. J. Immunol. 22:1803-1812. - PubMed
-
- Battegay, M., S. Cooper, A. Althage, J. Banziger, H. Hengartner, and R. M. Zinkernagel. 1991. Quantification of lymphocytic choriomeningitis virus with an immunological focus assay in 24- or 96-well plates. J. Virol. Methods 33:191-198. - PubMed
-
- Battegay, M., D. Moskophidis, H. Waldner, M. A. Brundler, W. P. Fung-Leung, T. W. Mak, H. Hengartner, and R. M. Zinkernagel. 1993. Impairment and delay of neutralizing antiviral antibody responses by virus-specific cytotoxic T cells. J. Immunol. 151:5408-5415. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
