

The topology of hepatitis B virus pregenomic RNA promotes its replication
- PMID: 17699570
- PMCID: PMC2168771
- DOI: 10.1128/JVI.01414-07
The topology of hepatitis B virus pregenomic RNA promotes its replication
Abstract
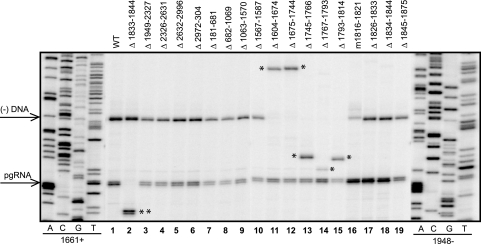
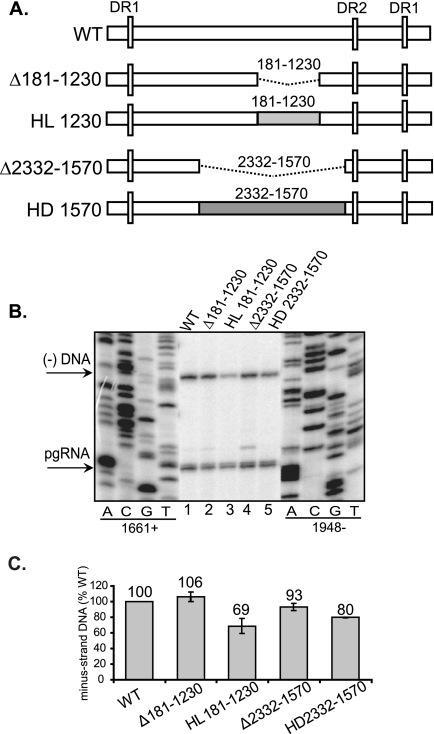
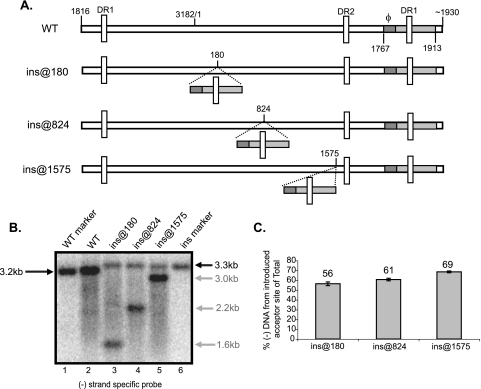
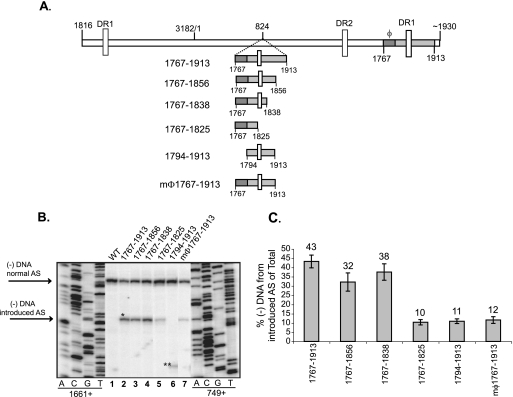
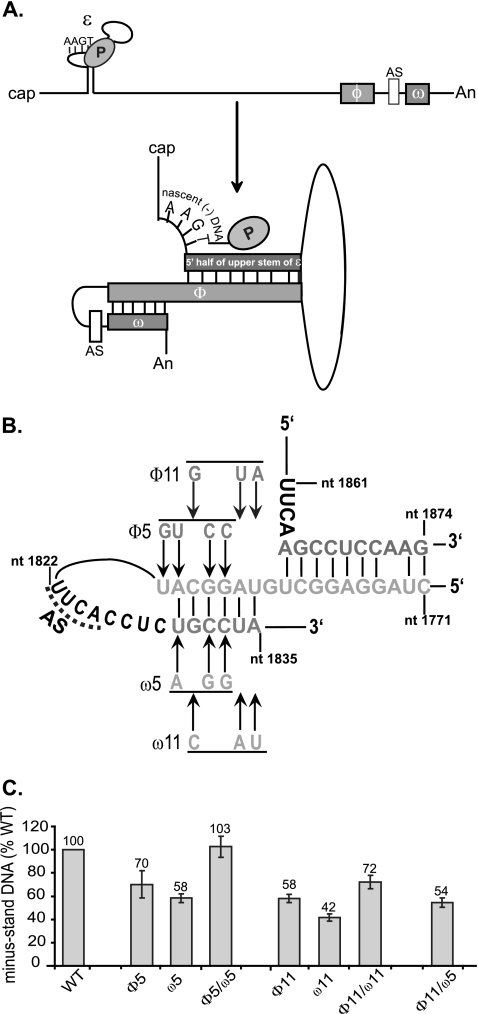
Previous analysis of hepatitis B virus (HBV) indicated base pairing between two cis-acting sequences, the 5' half of the upper stem of epsilon and phi, contributes to the synthesis of minus-strand DNA. Our goal was to identify other cis-acting sequences on the pregenomic RNA (pgRNA) involved in the synthesis of minus-strand DNA. We found that large portions of the pgRNA could be deleted or substituted without an appreciable decrease in the level of minus-strand DNA synthesized, indicating that most of the pgRNA is dispensable and that a specific size of the pgRNA is not required for this process. Our results indicated that the cis-acting sequences for the synthesis of minus-strand DNA are present near the 5' and 3' ends of the pgRNA. In addition, we found that the first-strand template switch could be directed to a new location when a 72-nucleotide (nt) fragment, which contained the cis-acting sequences present near the 3' end of the pgRNA, was introduced at that location. Within this 72-nt region, we uncovered two new cis-acting sequences, which flank the acceptor site. We show that one of these sequences, named omega and located 3' of the acceptor site, base pairs with phi to contribute to the synthesis of minus-strand DNA. Thus, base pairing between three cis-acting elements (5' half of the upper stem of epsilon, phi, and omega) are necessary for the synthesis of HBV minus-strand DNA. We propose that this topology of pgRNA facilitates first-strand template switch and/or the initiation of synthesis of minus-strand DNA.
Figures

References
-
- Geballe, A. P., R. R. Spaete, and E. S. Mocarski. 1986. A cis-acting element within the 5′ leader of a cytomegalovirus β transcript determines kinetic class. Cell 46:865-872. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Research Materials
