

Regulation of ubiquitin-proteasome system mediated degradation by cytosolic stress
- PMID: 17699585
- PMCID: PMC2043544
- DOI: 10.1091/mbc.e07-05-0487
Regulation of ubiquitin-proteasome system mediated degradation by cytosolic stress
Abstract
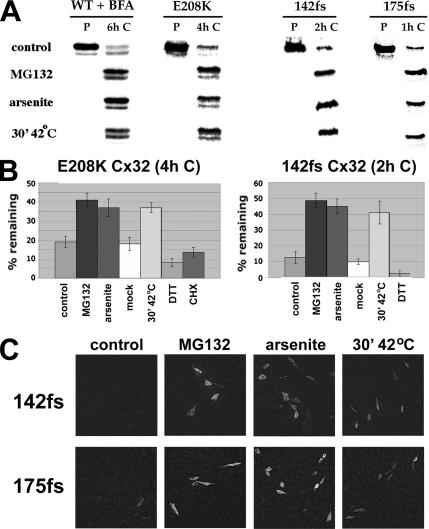
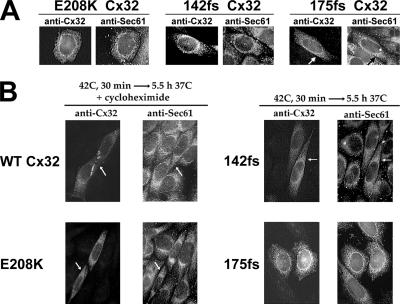
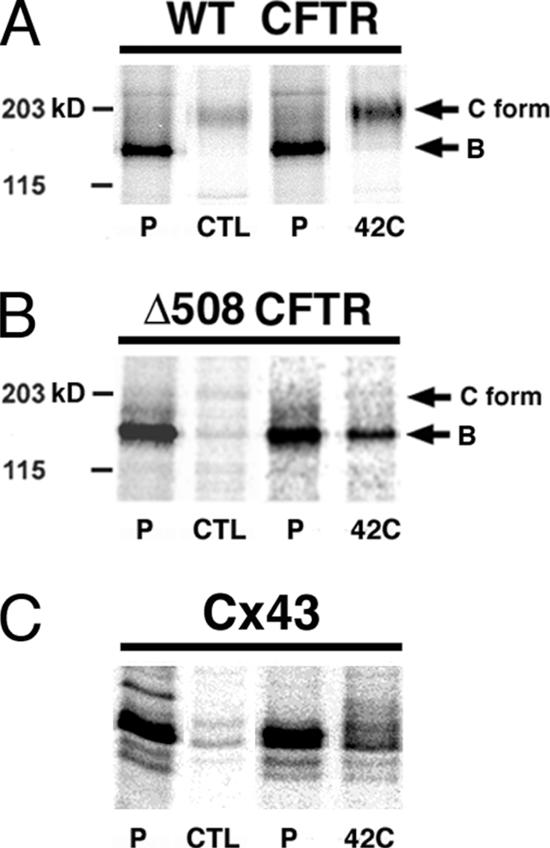
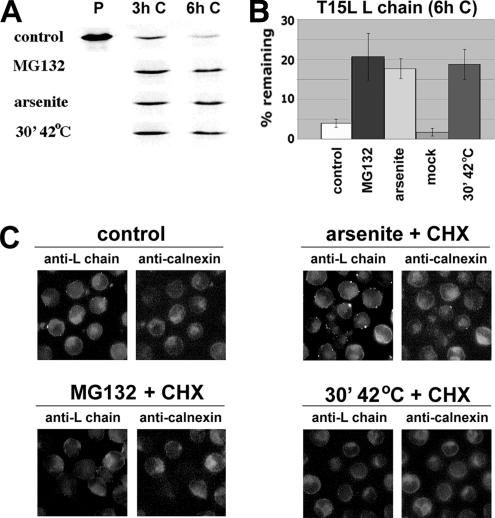
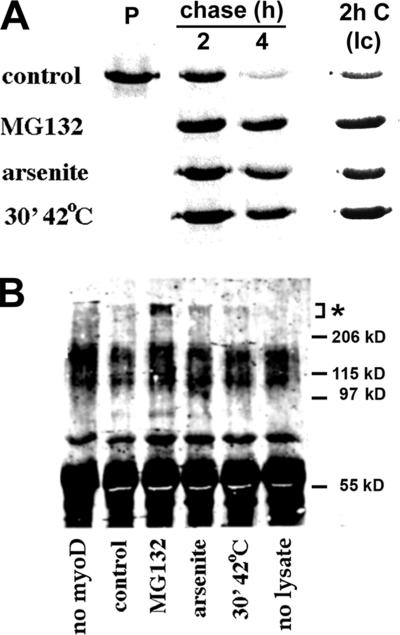
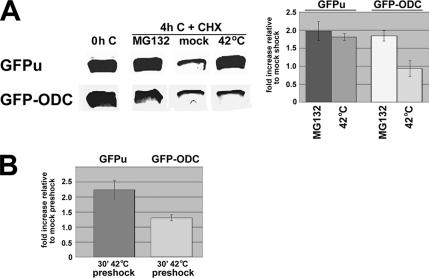
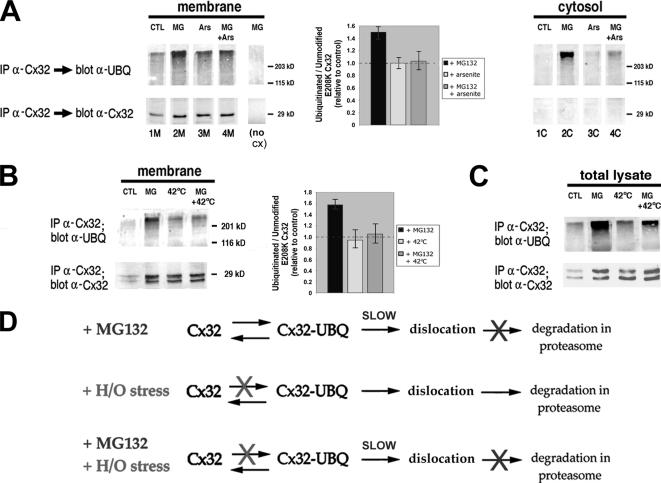
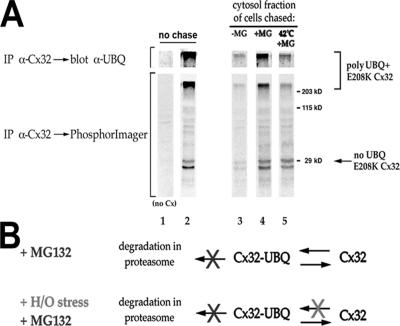
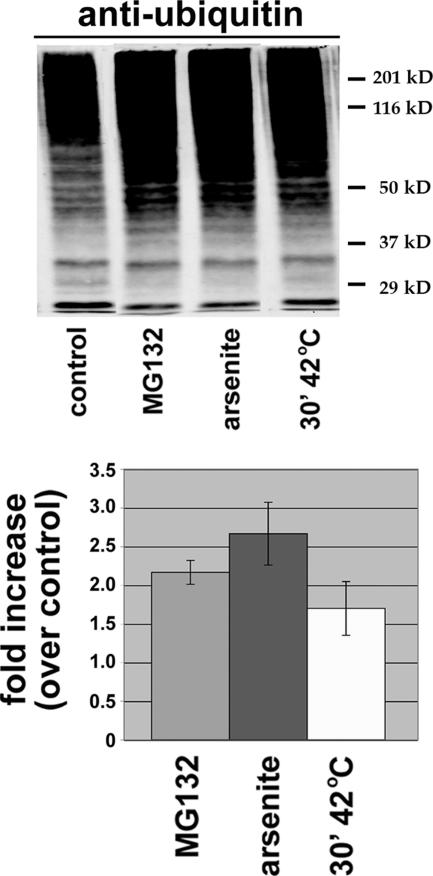
ER-associated, ubiquitin-proteasome system (UPS)-mediated degradation of the wild-type (WT) gap junction protein connexin32 (Cx32) is inhibited by mild forms of cytosolic stress at a step before its dislocation into the cytosol. We show that the same conditions (a 30-min, 42 degrees C heat shock or oxidative stress induced by arsenite) also reduce the endoplasmic reticulum (ER)-associated turnover of disease-causing mutants of Cx32 and the cystic fibrosis transmembrane conductance regulator (CFTR), as well as that of WT CFTR and unassembled Ig light chain. Stress-stabilized WT Cx32 and CFTR, but not the mutant/unassembled proteins examined, could traverse the secretory pathway. Heat shock also slowed the otherwise rapid UPS-mediated turnover of the cytosolic proteins myoD and GFPu, but not the degradation of an ubiquitination-independent construct (GFP-ODC) closely related to the latter. Analysis of mutant Cx32 from cells exposed to proteasome inhibitors and/or cytosolic stress indicated that stress reduces degradation at the level of substrate polyubiquitination. These findings reveal a new link between the cytosolic stress-induced heat shock response, ER-associated degradation, and polyubiquitination. Stress-denatured proteins may titer a limiting component of the ubiquitination machinery away from pre-existing UPS substrates, thereby sparing the latter from degradation.
Figures
Similar articles
-
Dislocation and degradation from the ER are regulated by cytosolic stress.J Cell Biol. 2002 Apr 29;157(3):381-94. doi: 10.1083/jcb.200111045. Epub 2002 Apr 29. J Cell Biol. 2002. PMID: 11980915 Free PMC article.
-
ER stress induces alternative nonproteasomal degradation of ER proteins but not of cytosolic ones.Cell Stress Chaperones. 2007 Winter;12(4):373-83. doi: 10.1379/csc-281.1. Cell Stress Chaperones. 2007. PMID: 18229456 Free PMC article.
-
Cytosolic stress reduces degradation of connexin43 internalized from the cell surface and enhances gap junction formation and function.Mol Biol Cell. 2005 Nov;16(11):5247-57. doi: 10.1091/mbc.e05-05-0415. Epub 2005 Aug 31. Mol Biol Cell. 2005. PMID: 16135529 Free PMC article.
-
Physiological and pathological role of the ubiquitin-proteasome system in the vascular smooth muscle cell.Cardiovasc Res. 2012 Jul 15;95(2):183-93. doi: 10.1093/cvr/cvs128. Epub 2012 Mar 26. Cardiovasc Res. 2012. PMID: 22451513 Review.
-
Role of the ubiquitin-proteasome system in brain ischemia: friend or foe?Prog Neurobiol. 2014 Jan;112:50-69. doi: 10.1016/j.pneurobio.2013.10.003. Epub 2013 Oct 22. Prog Neurobiol. 2014. PMID: 24157661 Review.
Cited by
-
Phosphorylation and activation of ubiquitin-specific protease-14 by Akt regulates the ubiquitin-proteasome system.Elife. 2015 Nov 2;4:e10510. doi: 10.7554/eLife.10510. Elife. 2015. PMID: 26523394 Free PMC article.
-
Connexin: a potential novel target for protecting the central nervous system?Neural Regen Res. 2015 Apr;10(4):659-66. doi: 10.4103/1673-5374.155444. Neural Regen Res. 2015. PMID: 26170830 Free PMC article. Review.
-
Proteasome inhibitor treatment in alcoholic liver disease.World J Gastroenterol. 2011 May 28;17(20):2558-62. doi: 10.3748/wjg.v17.i20.2558. World J Gastroenterol. 2011. PMID: 21633661 Free PMC article.
-
The cystic-fibrosis-associated ΔF508 mutation confers post-transcriptional destabilization on the C. elegans ABC transporter PGP-3.Dis Model Mech. 2012 Nov;5(6):930-9. doi: 10.1242/dmm.008987. Epub 2012 May 8. Dis Model Mech. 2012. PMID: 22569626 Free PMC article.
-
TRIM11 activates the proteasome and promotes overall protein degradation by regulating USP14.Nat Commun. 2018 Mar 26;9(1):1223. doi: 10.1038/s41467-018-03499-z. Nat Commun. 2018. PMID: 29581427 Free PMC article.
References
-
- Abel A., Bone L. J., Messing A., Scherer S. S., Fischbeck K. H. Studies in transgenic mice indicate a loss of connexin32 function in X-linked Charcot-Marie-Tooth disease. J. Neuropathol. Exp. Neurol. 1999;58:702–710. - PubMed
-
- Bence N. F., Bennett E. J., Kopito R. R. Application and analysis of the GFPu family of ubiquitin-proteasome system reporters. Methods Enzymol. 2005;399:481–490. - PubMed
-
- Bond U., Agell N., Haas A. L., Redman K., Schlesinger M. J. Ubiquitin in stressed chicken embryo fibroblasts. J. Biol. Chem. 1988;263:2384–2388. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Research Materials
Miscellaneous
