

Atg18 regulates organelle morphology and Fab1 kinase activity independent of its membrane recruitment by phosphatidylinositol 3,5-bisphosphate
- PMID: 17699591
- PMCID: PMC2043547
- DOI: 10.1091/mbc.e07-04-0301
Atg18 regulates organelle morphology and Fab1 kinase activity independent of its membrane recruitment by phosphatidylinositol 3,5-bisphosphate
Abstract
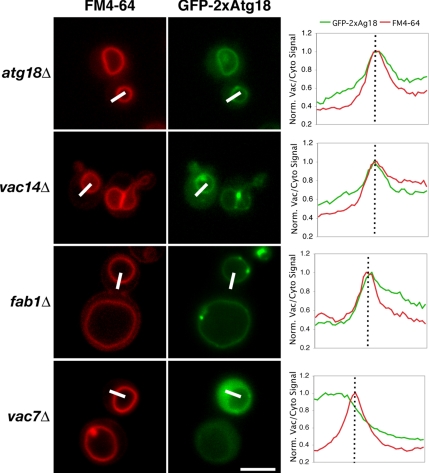
The lipid kinase Fab1 governs yeast vacuole homeostasis by generating PtdIns(3,5)P(2) on the vacuolar membrane. Recruitment of effector proteins by the phospholipid ensures precise regulation of vacuole morphology and function. Cells lacking the effector Atg18p have enlarged vacuoles and high PtdIns(3,5)P(2) levels. Although Atg18 colocalizes with Fab1p, it likely does not directly interact with Fab1p, as deletion of either kinase activator-VAC7 or VAC14-is epistatic to atg18Delta: atg18Deltavac7Delta cells have no detectable PtdIns(3,5)P(2). Moreover, a 2xAtg18 (tandem fusion) construct localizes to the vacuole membrane in the absence of PtdIns(3,5)P(2), but requires Vac7p for recruitment. Like the endosomal PtdIns(3)P effector EEA1, Atg18 membrane binding may require a protein component. When the lipid requirement is bypassed by fusing Atg18 to ALP, a vacuolar transmembrane protein, vac14Delta vacuoles regain normal morphology. Rescue is independent of PtdIns(3,5)P(2), as mutation of the phospholipid-binding site in Atg18 does not prevent vacuole fission and properly regulates Fab1p activity. Finally, the vacuole-specific type-V myosin adapter Vac17p interacts with Atg18p, perhaps mediating cytoskeletal attachment during retrograde transport. Atg18p is likely a PtdIns(3,5)P(2)"sensor," acting as an effector to remodel membranes as well as regulating its synthesis via feedback that might involve Vac7p.
Figures






References
-
- Balla T. Inositol-lipid binding motifs: signal integrators through protein-lipid and protein-protein interactions. J. Cell Sci. 2005;118:2093–2104. - PubMed
-
- Barth H., Meiling-Wesse K., Epple U. D., Thumm M. Autophagy and the cytoplasm to vacuole targeting pathway both require Aut10p. FEBS Lett. 2001;508:23–28. - PubMed
-
- Behnia R., Munro S. Organelle identity and the signposts for membrane traffic. Nature. 2005;438:597–604. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
