

Neuronal diversity in GABAergic long-range projections from the hippocampus
- PMID: 17699661
- PMCID: PMC2270609
- DOI: 10.1523/JNEUROSCI.1847-07.2007
Neuronal diversity in GABAergic long-range projections from the hippocampus
Abstract
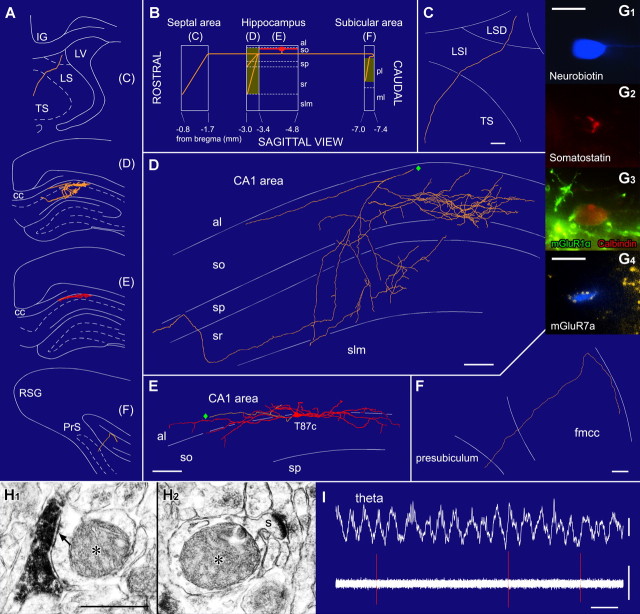
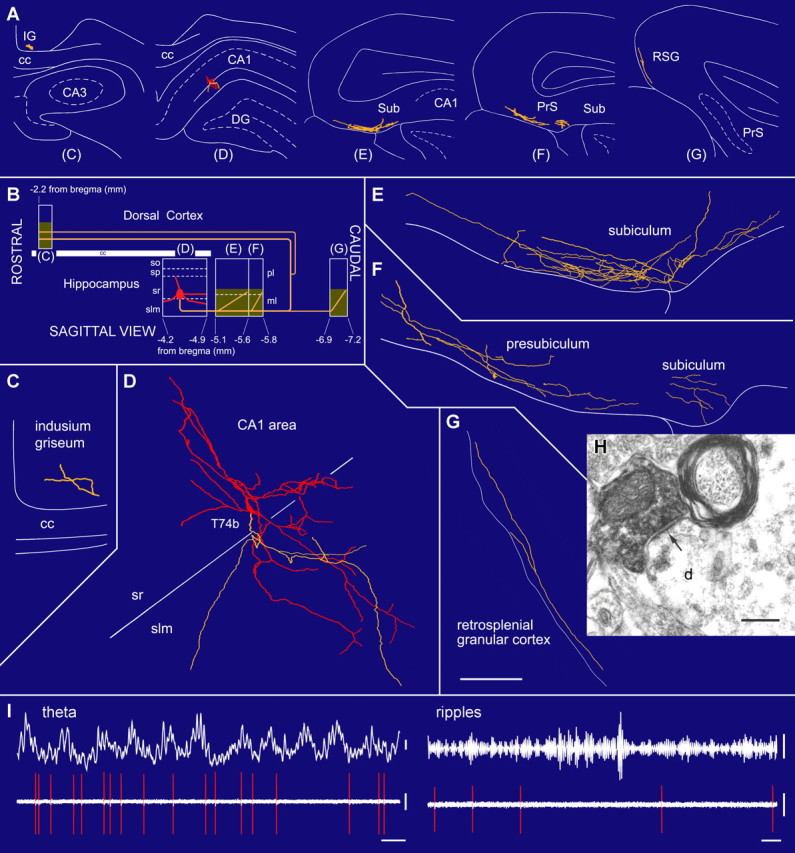
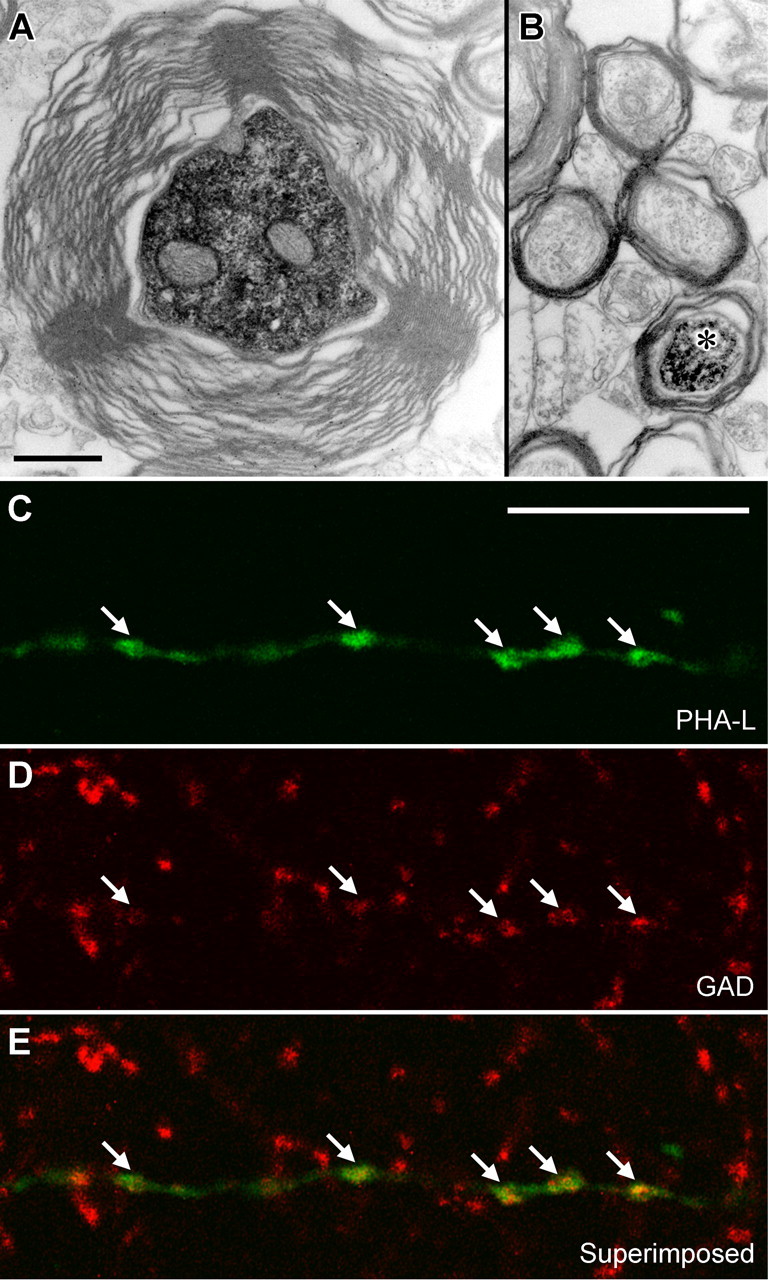
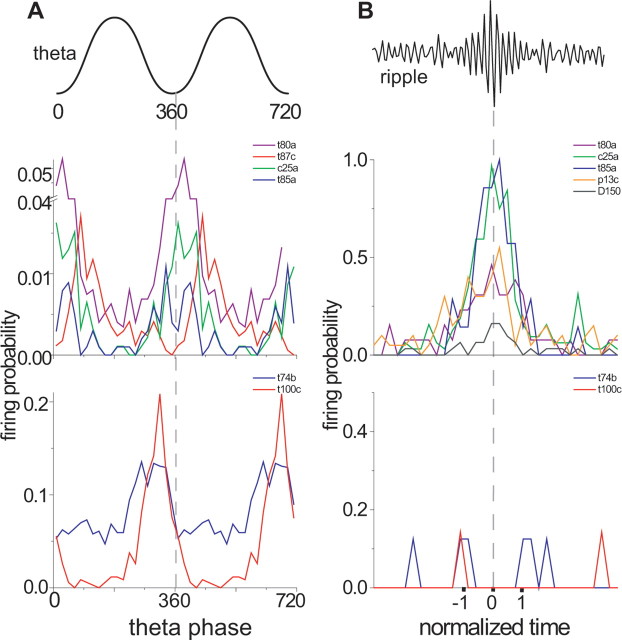
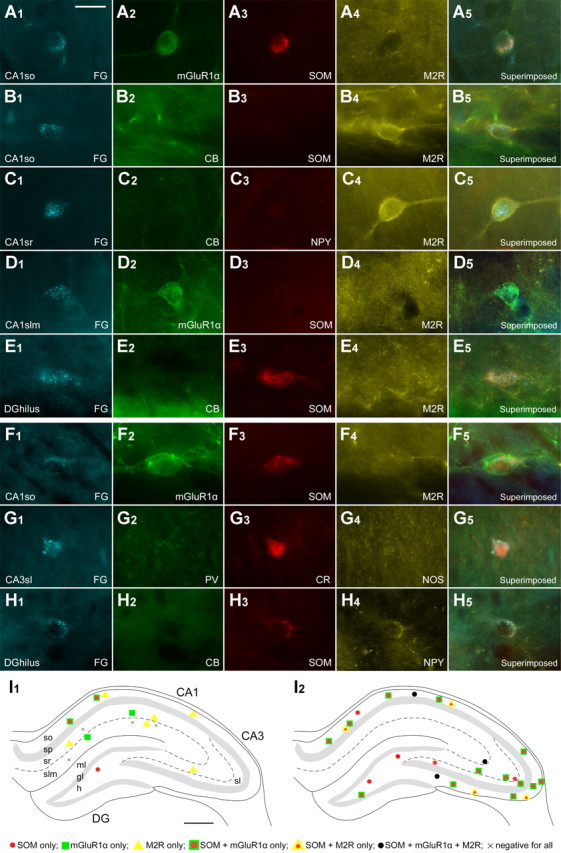
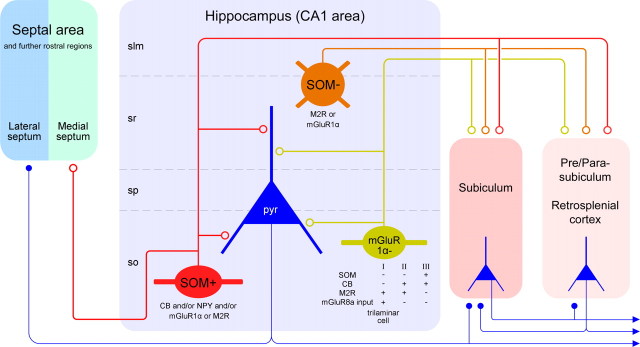
The formation and recall of sensory, motor, and cognitive representations require coordinated fast communication among multiple cortical areas. Interareal projections are mainly mediated by glutamatergic pyramidal cell projections; only few long-range GABAergic connections have been reported. Using in vivo recording and labeling of single cells and retrograde axonal tracing, we demonstrate novel long-range GABAergic projection neurons in the rat hippocampus: (1) somatostatin- and predominantly mGluR1alpha-positive neurons in stratum oriens project to the subiculum, other cortical areas, and the medial septum; (2) neurons in stratum oriens, including somatostatin-negative ones; and (3) trilaminar cells project to the subiculum and/or other cortical areas but not the septum. These three populations strongly increase their firing during sharp wave-associated ripple oscillations, communicating this network state to the septotemporal system. Finally, a large population of somatostatin-negative GABAergic cells in stratum radiatum project to the molecular layers of the subiculum, presubiculum, retrosplenial cortex, and indusium griseum and fire rhythmically at high rates during theta oscillations but do not increase their firing during ripples. The GABAergic projection axons have a larger diameter and thicker myelin sheet than those of CA1 pyramidal cells. Therefore, rhythmic IPSCs are likely to precede the arrival of excitation in cortical areas (e.g., subiculum) that receive both glutamatergic and GABAergic projections from the CA1 area. Other areas, including the retrosplenial cortex, receive only rhythmic GABAergic CA1 input. We conclude that direct GABAergic projections from the hippocampus to other cortical areas and the septum contribute to coordinating oscillatory timing across structures.
Figures

Similar articles
-
GABAergic projections from the hippocampus to the retrosplenial cortex in the rat.Eur J Neurosci. 2007 Sep;26(5):1193-204. doi: 10.1111/j.1460-9568.2007.05745.x. Eur J Neurosci. 2007. PMID: 17767498
-
Behavior-dependent activity patterns of GABAergic long-range projecting neurons in the rat hippocampus.Hippocampus. 2017 Apr;27(4):359-377. doi: 10.1002/hipo.22696. Epub 2017 Feb 2. Hippocampus. 2017. PMID: 27997999 Free PMC article.
-
Spatio-temporal specialization of GABAergic septo-hippocampal neurons for rhythmic network activity.Brain Struct Funct. 2018 Jun;223(5):2409-2432. doi: 10.1007/s00429-018-1626-0. Epub 2018 Mar 3. Brain Struct Funct. 2018. PMID: 29500537 Free PMC article.
-
Spatial organization of direct hippocampal field CA1 axonal projections to the rest of the cerebral cortex.Brain Res Rev. 2007 Nov;56(1):1-26. doi: 10.1016/j.brainresrev.2007.05.002. Epub 2007 May 10. Brain Res Rev. 2007. PMID: 17559940 Free PMC article. Review.
-
Hippocampus as comparator: role of the two input and two output systems of the hippocampus in selection and registration of information.Hippocampus. 2001;11(5):578-98. doi: 10.1002/hipo.1073. Hippocampus. 2001. PMID: 11732710 Review.
Cited by
-
Molecular mechanism of circadian rhythmicity of seizures in temporal lobe epilepsy.Front Cell Neurosci. 2012 Nov 23;6:55. doi: 10.3389/fncel.2012.00055. eCollection 2012. Front Cell Neurosci. 2012. PMID: 23189039 Free PMC article.
-
Hippocampal efferents to retrosplenial cortex and lateral septum are required for memory acquisition.Learn Mem. 2020 Jul 15;27(8):310-318. doi: 10.1101/lm.051797.120. Print 2020 Aug. Learn Mem. 2020. PMID: 32669386 Free PMC article.
-
Fast gamma oscillations are generated intrinsically in CA1 without the involvement of fast-spiking basket cells.J Neurosci. 2015 Feb 25;35(8):3616-24. doi: 10.1523/JNEUROSCI.4166-14.2015. J Neurosci. 2015. PMID: 25716860 Free PMC article.
-
Inhibition is a prevalent mode of activity in the neocortex around awake hippocampal ripples in mice.Elife. 2023 Jan 16;12:e79513. doi: 10.7554/eLife.79513. Elife. 2023. PMID: 36645126 Free PMC article.
-
Subcortical control of the default mode network: Role of the basal forebrain and implications for neuropsychiatric disorders.Brain Res Bull. 2022 Jul;185:129-139. doi: 10.1016/j.brainresbull.2022.05.005. Epub 2022 May 11. Brain Res Bull. 2022. PMID: 35562013 Free PMC article. Review.
References
-
- Acsady L, Gorcs TJ, Freund TF. Different populations of vasoactive intestinal polypeptide-immunoreactive interneurons are specialized to control pyramidal cells or interneurons in the hippocampus. Neuroscience. 1996;73:317–334. - PubMed
-
- Alonso A, Kohler C. Evidence for separate projections of hippocampal pyramidal and non-pyramidal neurons to different parts of the septum in the rat brain. Neurosci Lett. 1982;31:209–214. - PubMed
-
- Amaral DG, Witter MP. The three-dimensional organization of the hippocampal formation: a review of anatomical data. Neuroscience. 1989;31:571–591. - PubMed
-
- Bibbig A, Traub RD, Whittington MA. Long-range synchronization of γ and β oscillations and the plasticity of excitatory and inhibitory synapses: a network model. J Neurophysiol. 2002;88:1634–1654. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Miscellaneous