

Modulation of thalamic nociceptive processing after spinal cord injury through remote activation of thalamic microglia by cysteine cysteine chemokine ligand 21
- PMID: 17699671
- PMCID: PMC6672166
- DOI: 10.1523/JNEUROSCI.2209-07.2007
Modulation of thalamic nociceptive processing after spinal cord injury through remote activation of thalamic microglia by cysteine cysteine chemokine ligand 21
Abstract
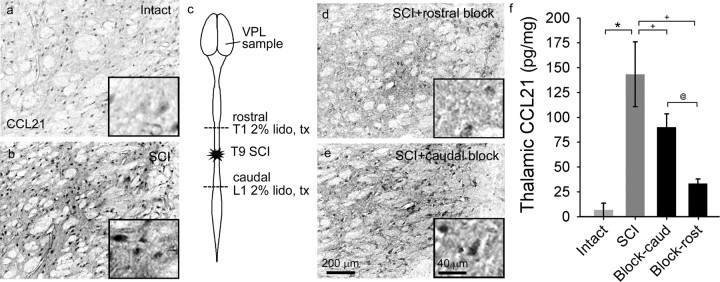
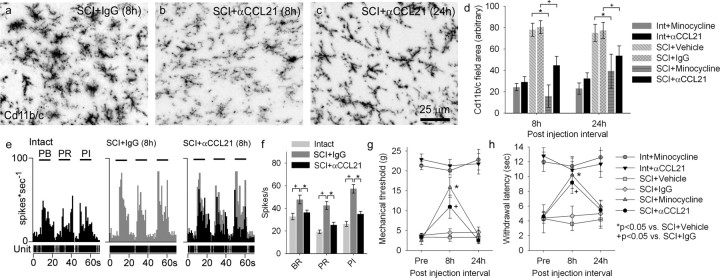
Spinal cord injury (SCI) results in the generation and amplification of pain caused in part by injury-induced changes in neuronal excitability at multiple levels along the sensory neuraxis. We have previously shown that activated microglia, through an ERK (extracellular signal-regulated kinase)-regulated PGE(2) (prostaglandin E(2)) signaling mechanism, maintain neuronal hyperexcitability in the lumbar dorsal horn. Here, we examined whether microglial cells in the thalamus contribute to the modulation of chronic pain after SCI, and whether microglial activation is governed by spinally mediated increases in the microglial activator cysteine-cysteine chemokine ligand 21 (CCL21). We report that CCL21 is upregulated in dorsal horn neurons, that tissue levels are increased in the dorsal horn and ventral posterolateral (VPL) nucleus of the thalamus 4 weeks after SCI, and that the increase can be differentially reduced by spinal blockade at T1 or L1. In intact animals, electrical stimulation of the spinothalamic tract induces increases in thalamic CCL21 levels. Recombinant CCL21 injected into the VPL of intact animals transiently activates microglia and induces pain-related behaviors, effects that could be blocked with minocycline. After SCI, intra-VPL antibody-mediated neutralization of CCL21 decreases microglial activation and evoked hyperexcitability of VPL neurons, and restores nociceptive thresholds to near-normal levels. These data identify a novel pathway by which SCI triggers upregulation of the neuroimmune modulator CCL21 in the thalamus, which induces microglial activation in association with pain phenomena.
Figures




Similar articles
-
Cell cycle activation contributes to increased neuronal activity in the posterior thalamic nucleus and associated chronic hyperesthesia after rat spinal cord contusion.Neurotherapeutics. 2013 Jul;10(3):520-38. doi: 10.1007/s13311-013-0198-1. Neurotherapeutics. 2013. PMID: 23775067 Free PMC article.
-
Extracellular signal-regulated kinase-regulated microglia-neuron signaling by prostaglandin E2 contributes to pain after spinal cord injury.J Neurosci. 2007 Feb 28;27(9):2357-68. doi: 10.1523/JNEUROSCI.0138-07.2007. J Neurosci. 2007. PMID: 17329433 Free PMC article.
-
Estradiol attenuates spinal cord injury-induced pain by suppressing microglial activation in thalamic VPL nuclei of rats.Neurosci Res. 2013 Apr;75(4):316-23. doi: 10.1016/j.neures.2013.01.010. Epub 2013 Feb 16. Neurosci Res. 2013. PMID: 23419864
-
Sodium channel expression and the molecular pathophysiology of pain after SCI.Prog Brain Res. 2007;161:195-203. doi: 10.1016/S0079-6123(06)61013-3. Prog Brain Res. 2007. PMID: 17618978 Review.
-
Fire and phantoms after spinal cord injury: Na+ channels and central pain.Trends Neurosci. 2006 Apr;29(4):207-15. doi: 10.1016/j.tins.2006.02.003. Epub 2006 Feb 21. Trends Neurosci. 2006. PMID: 16494954 Review.
Cited by
-
Delayed microglial depletion after spinal cord injury reduces chronic inflammation and neurodegeneration in the brain and improves neurological recovery in male mice.Theranostics. 2020 Sep 14;10(25):11376-11403. doi: 10.7150/thno.49199. eCollection 2020. Theranostics. 2020. PMID: 33052221 Free PMC article.
-
Endoplasmic Reticulum Stress and Disrupted Neurogenesis in the Brain Are Associated with Cognitive Impairment and Depressive-Like Behavior after Spinal Cord Injury.J Neurotrauma. 2016 Nov 1;33(21):1919-1935. doi: 10.1089/neu.2015.4348. Epub 2016 May 16. J Neurotrauma. 2016. PMID: 27050417 Free PMC article.
-
Pathological activity in mediodorsal thalamus of rats with spinal cord injury pain.J Neurosci. 2013 Feb 27;33(9):3915-26. doi: 10.1523/JNEUROSCI.2639-12.2013. J Neurosci. 2013. PMID: 23447602 Free PMC article.
-
Neuroinflammatory responses of microglia in central nervous system trauma.J Cereb Blood Flow Metab. 2020 Dec;40(1_suppl):S25-S33. doi: 10.1177/0271678X20965786. Epub 2020 Oct 22. J Cereb Blood Flow Metab. 2020. PMID: 33086921 Free PMC article. Review.
-
Spinal glia modulate both adaptive and pathological processes.Brain Behav Immun. 2009 Oct;23(7):969-76. doi: 10.1016/j.bbi.2009.05.001. Epub 2009 May 10. Brain Behav Immun. 2009. PMID: 19435601 Free PMC article.
References
-
- Bernards CM. Epidural and intrathecal drug movement. In: Yaksh TL, editor. Spinal drug delivery. Amsterdam: Elsevier; 1999. pp. 239–252.
-
- Biber K, Sauter A, Brouwer N, Copray SC, Boddeke HW. Ischemia-induced neuronal expression of the microglia attracting chemokine secondary lymphoid-tissue chemokine (SLC) Glia. 2001;34:121–133. - PubMed
-
- Biber K, Dijkstra I, Trebst C, De Groot CJA, Ransohoff RM, Boddeke HWGM. Functional expression of CXCR3 in cultured mouse and human astrocytes and microglia. Neuroscience. 2002;112:487–497. - PubMed
-
- Chang HT. Subacute human spinal cord contusion: few lymphocytes and many macrophages. Spinal Cord. 2007;45:174–182. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Medical
Miscellaneous