

Protein/DNA arrays identify nitric oxide-regulated cis-element and trans-factor activities some of which govern neuroblastoma cell viability
- PMID: 17702766
- PMCID: PMC2018649
- DOI: 10.1093/nar/gkm594
Protein/DNA arrays identify nitric oxide-regulated cis-element and trans-factor activities some of which govern neuroblastoma cell viability
Abstract
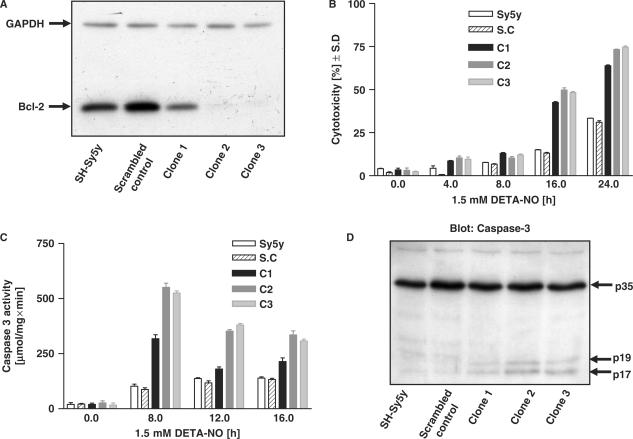
Toxic nitric oxide (NO) levels can regulate gene expression. Using a novel protein/DNA array, we show that toxic NO levels regulate the binding of trans-factors to various cis-elements in neuroblastoma cells, including CRE and those recognized by the transcription factors AP1, AP2, Brn-3a, EGR, E2F1 and SP1. Functionality of some of the cis-elements was confirmed by electro mobility shift and reporter assays. Interestingly, CREB, AP-1, Brn-3a, EGR and E2F1 can control mammalian cell viability. NO induced the anti-apoptotic Bcl-2 protein and its mRNA prior to the onset of death of 30-60% of the cells. Promoter analysis of the bcl-2 gene confirmed the involvement of a CRE in NO-dependent bcl-2 transcription. Neuroblastoma cells over-expressing bcl-2 became much more resistant to NO-induced apoptosis; conversely, Bcl-2 knockdown cells were rendered markedly more sensitive to NO. Together these results suggest that Bcl-2 counteracts NO-induced apoptosis in a fraction of the cell population. Thus, NO stimulates the binding of many trans-factors to their cognate cis-elements, some of which can regulate cell viability through transcriptional activation of target genes. Our results emphasize that a DNA/protein array approach can reveal novel, global transcription factor activities stimulated by cell death-regulating molecules.
Figures







References
-
- Hemish J, Nakaya N, Mittal V, Enikolopov G. Nitric oxide activates diverse signaling pathways to regulate gene expression. J. Biol. Chem. 2003;278:42321–42329. - PubMed
-
- Moncada S, Erusalimsky JD. Does nitric oxide modulate mitochondrial energy generation and apoptosis? Nat. Rev. Mol. Cell. Biol. 2002;3:214–220. - PubMed
-
- Lucas KA, Pitari GM, Kazerounian S, Ruiz-Stewart I, Park J, Schulz S, Chepenik KP, Waldman SA. Guanylyl cyclases and signaling by cyclic GMP. Pharmacol. Rev. 2000;52:375–414. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Research Materials
