

The delta2 'ionotropic' glutamate receptor functions as a non-ionotropic receptor to control cerebellar synaptic plasticity
- PMID: 17702810
- PMCID: PMC2277055
- DOI: 10.1113/jphysiol.2007.141291
The delta2 'ionotropic' glutamate receptor functions as a non-ionotropic receptor to control cerebellar synaptic plasticity
Abstract
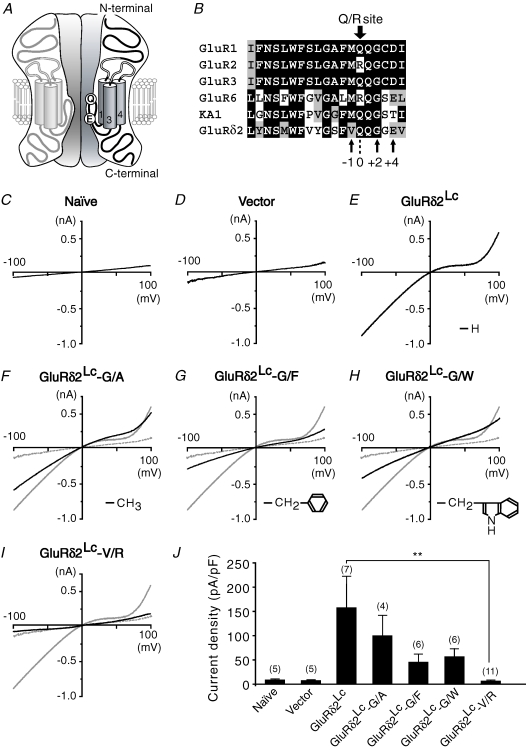
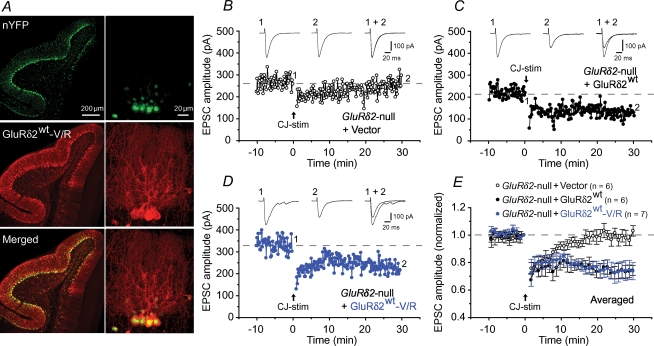
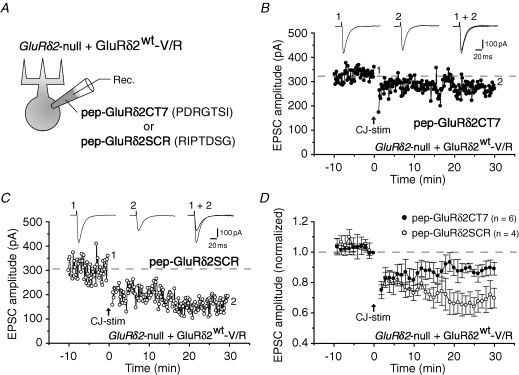
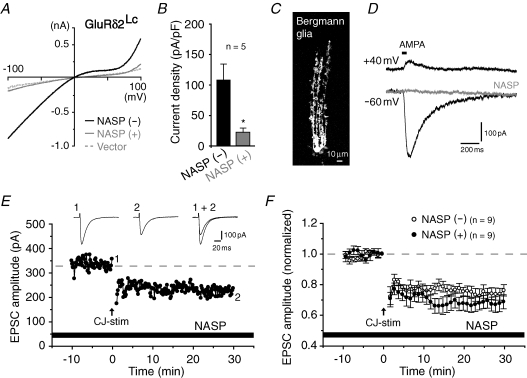
The delta2 glutamate receptor (GluRdelta2) belongs to the ionotropic glutamate receptor (iGluR) family and plays a crucial role in the induction of cerebellar long-term depression (LTD), a form of synaptic plasticity underlying motor learning. Nevertheless, the mechanisms by which GluRdelta2 regulates cerebellar LTD have remained elusive. Because a mutation occurring in lurcher mice causes continuous GluRdelta2 channel activity that can be abolished by 1-naphtylacetylspermine (NASP), a channel blocker for Ca(2+)-permeable iGluRs, GluRdelta2 is thought to function as an ion channel. Here, we introduced a mutant GluRdelta2 transgene, in which the putative channel pore was disrupted, into GluRdelta2-null Purkinje cells using a virus vector. Surprisingly and similar to the effect of the wild-type GluRdelta2 transgene, the mutant GluRdelta2 completely rescued the abrogated LTD in GluRdelta2-null mice. Furthermore, NASP did not block LTD induction in wild-type cerebellar slices. These results indicate that GluRdelta2, a member of the iGluR family, does not serve as a channel in the regulation of LTD induction.
Figures
Similar articles
-
Ca2+ permeability of the channel pore is not essential for the delta2 glutamate receptor to regulate synaptic plasticity and motor coordination.J Physiol. 2007 Mar 15;579(Pt 3):729-35. doi: 10.1113/jphysiol.2006.127100. Epub 2007 Jan 25. J Physiol. 2007. PMID: 17255161 Free PMC article.
-
The extreme C-terminus of GluRdelta2 is essential for induction of long-term depression in cerebellar slices.Eur J Neurosci. 2007 Mar;25(5):1357-62. doi: 10.1111/j.1460-9568.2007.05412.x. Eur J Neurosci. 2007. PMID: 17425562
-
Induction of long-term depression and phosphorylation of the delta2 glutamate receptor by protein kinase C in cerebellar slices.Eur J Neurosci. 2005 Oct;22(7):1817-20. doi: 10.1111/j.1460-9568.2005.04319.x. Eur J Neurosci. 2005. PMID: 16197524
-
The delta2 glutamate receptor: 10 years later.Neurosci Res. 2003 May;46(1):11-22. doi: 10.1016/s0168-0102(03)00036-1. Neurosci Res. 2003. PMID: 12725908 Review.
-
The delta2 glutamate receptor: a key molecule controlling synaptic plasticity and structure in Purkinje cells.Cerebellum. 2004;3(2):89-93. doi: 10.1080/14734220410028921. Cerebellum. 2004. PMID: 15233575 Review.
Cited by
-
The δ2 glutamate receptor gates long-term depression by coordinating interactions between two AMPA receptor phosphorylation sites.Proc Natl Acad Sci U S A. 2013 Mar 5;110(10):E948-57. doi: 10.1073/pnas.1218380110. Epub 2013 Feb 19. Proc Natl Acad Sci U S A. 2013. PMID: 23431139 Free PMC article.
-
Unlocking the secrets of the δ2 glutamate receptor: A gatekeeper for synaptic plasticity in the cerebellum.Commun Integr Biol. 2013 Nov 1;6(6):e26466. doi: 10.4161/cib.26466. Epub 2013 Sep 27. Commun Integr Biol. 2013. PMID: 24563706 Free PMC article.
-
The glutamate receptor subunit delta2 is capable of gating its intrinsic ion channel as revealed by ligand binding domain transplantation.Proc Natl Acad Sci U S A. 2009 Jun 23;106(25):10320-5. doi: 10.1073/pnas.0900329106. Epub 2009 Jun 8. Proc Natl Acad Sci U S A. 2009. PMID: 19506248 Free PMC article.
-
Structural basis for integration of GluD receptors within synaptic organizer complexes.Science. 2016 Jul 15;353(6296):295-9. doi: 10.1126/science.aae0104. Science. 2016. PMID: 27418511 Free PMC article.
-
Lack of evidence for direct ligand-gated ion channel activity of GluD receptors.Proc Natl Acad Sci U S A. 2024 Jul 30;121(31):e2406655121. doi: 10.1073/pnas.2406655121. Epub 2024 Jul 25. Proc Natl Acad Sci U S A. 2024. PMID: 39052831 Free PMC article.
References
-
- Araki K, Meguro H, Kushiya E, Takayama C, Inoue Y, Mishina M. Selective expression of the glutamate receptor channel δ2 subunit in cerebellar Purkinje cells. Biochem Biophys Res Commun. 1993;197:1267–1276. - PubMed
-
- Bowie D, Mayer ML. Inward rectification of both AMPA and kainate subtype glutamate receptors generated by polyamine-mediated ion channel block. Neuron. 1995;15:453–462. - PubMed
-
- Burnashev N, Monyer H, Seeburg PH, Sakmann B. Divalent ion permeability of AMPA receptor channels is dominated by the edited form of a single subunit. Neuron. 1992;8:189–198. - PubMed
-
- Hayashi T, Umemori H, Mishina M, Yamamoto T. The AMPA receptor interacts with and signals through the protein tyrosine kinase Lyn. Nature. 1999;397:72–76. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Miscellaneous