

Regionalization of the mouse visceral endoderm as the blastocyst transforms into the egg cylinder
- PMID: 17705827
- PMCID: PMC1978209
- DOI: 10.1186/1471-213X-7-96
Regionalization of the mouse visceral endoderm as the blastocyst transforms into the egg cylinder
Abstract
Background: Reciprocal interactions between two extra-embryonic tissues, the extra-embryonic ectoderm and the visceral endoderm, and the pluripotent epiblast, are required for the establishment of anterior-posterior polarity in the mouse. After implantation, two visceral endoderm cell types can be distinguished, in the embryonic and extra-embryonic regions of the egg cylinder. In the embryonic region, the specification of the anterior visceral endoderm (AVE) is central to the process of anterior-posterior patterning. Despite recent advances in our understanding of the molecular interactions underlying the differentiation of the visceral endoderm, little is known about how cells colonise the three regions of the tissue.
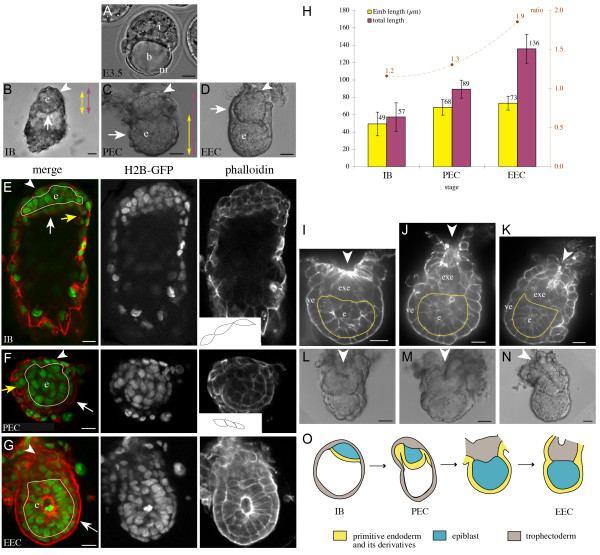
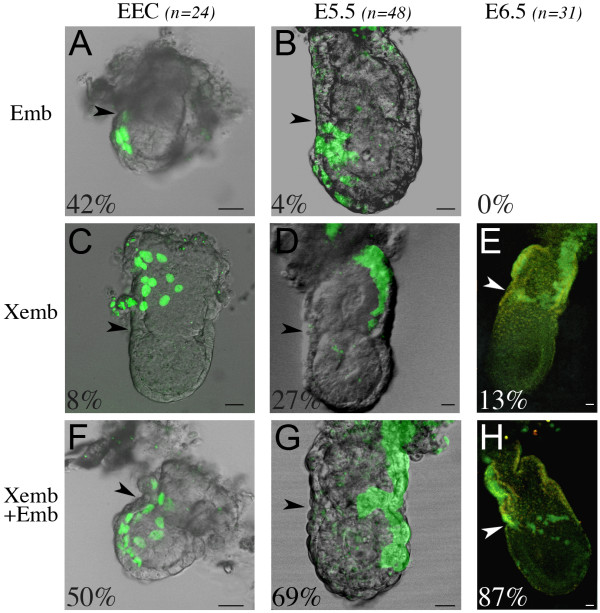
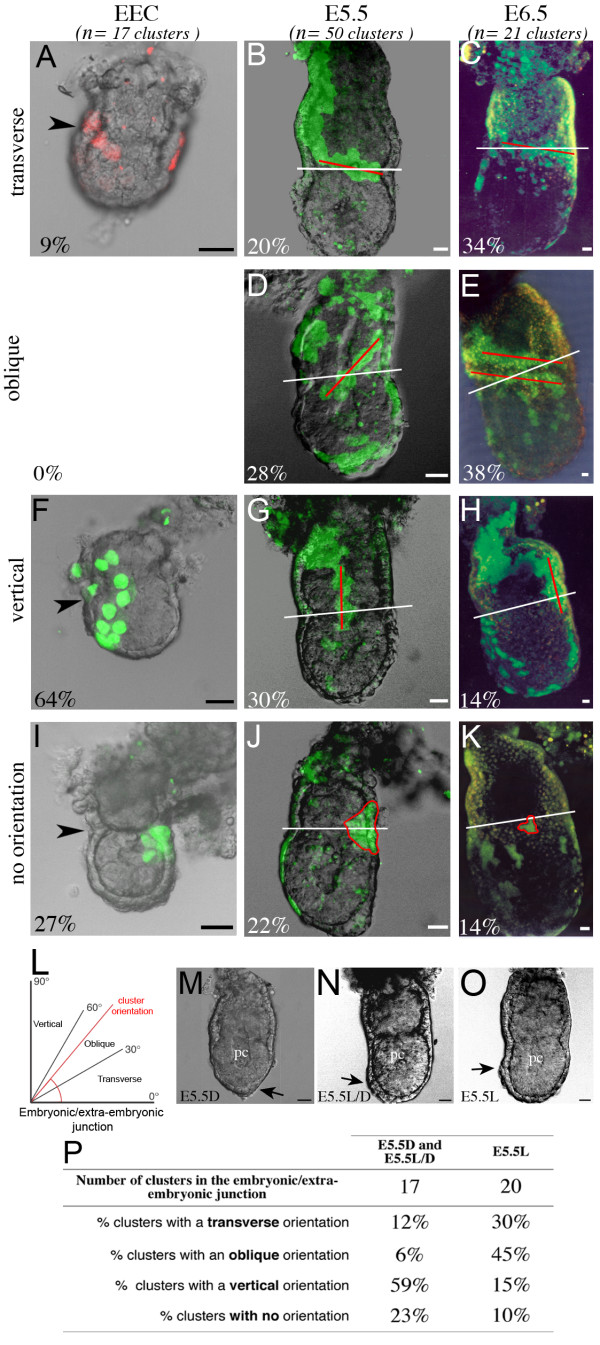
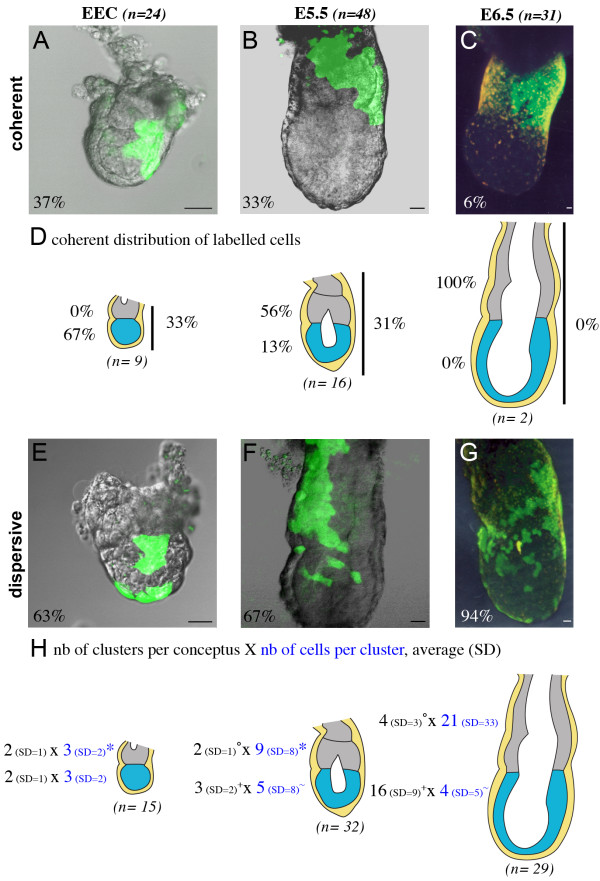
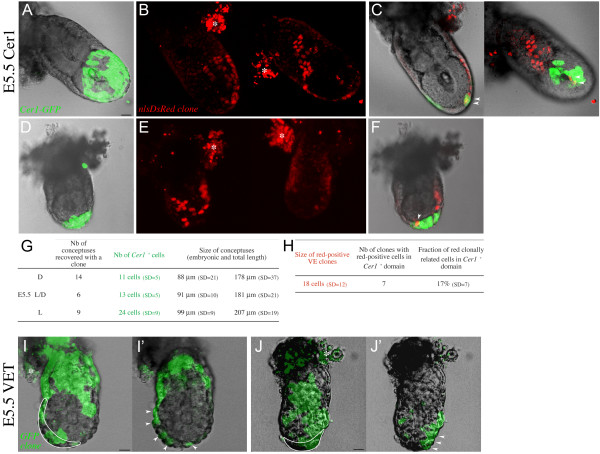
Results: As a first step, we performed morphological observations to understand how the extra-embryonic region of the egg cylinder forms from the blastocyst. Our analysis suggests a new model for the formation of this region involving cell rearrangements such as folding of the extra-embryonic ectoderm at the early egg cylinder stage. To trace visceral endoderm cells, we microinjected mRNAs encoding fluorescent proteins into single surface cells of the inner cell mass of the blastocyst and analysed the distribution of labelled cells at E5.0, E5.5 and E6.5. We found that at E5.0 the embryonic and extra-embryonic regions of the visceral endoderm do not correspond to distinct cellular compartments. Clusters of labelled cells may span the junction between the two regions even after the appearance of histological and molecular differences at E5.5. We show that in the embryonic region cell dispersion increases after the migration of the AVE. At this time, visceral endoderm cell clusters tend to become oriented parallel to the junction between the embryonic and extra-embryonic regions. Finally we investigated the origin of the AVE and demonstrated that this anterior signalling centre arises from more than a single precursor between E3.5 and E5.5.
Conclusion: We propose a new model for the formation of the extra-embryonic region of the egg cylinder involving a folding of the extra-embryonic ectoderm. Our analyses of the pattern of labelled visceral endoderm cells indicate that distinct cell behaviour in the embryonic and extra-embryonic regions is most apparent upon AVE migration. We also demonstrate the polyclonal origin of the AVE. Taken together, these studies lead to further insights into the formation of the extra-embryonic tissues as they first develop after implantation.
Figures
References
-
- Copp AJ. Interaction between inner cell mass and trophectoderm of the mouse blastocyst. II. The fate of the polar trophectoderm. J Embryol Exp Morphol. 1979;51:109–120. - PubMed
Publication types
MeSH terms
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Research Materials
