

JBP2, a SWI2/SNF2-like protein, regulates de novo telomeric DNA glycosylation in bloodstream form Trypanosoma brucei
- PMID: 17706299
- PMCID: PMC4735730
- DOI: 10.1016/j.molbiopara.2007.06.010
JBP2, a SWI2/SNF2-like protein, regulates de novo telomeric DNA glycosylation in bloodstream form Trypanosoma brucei
Abstract
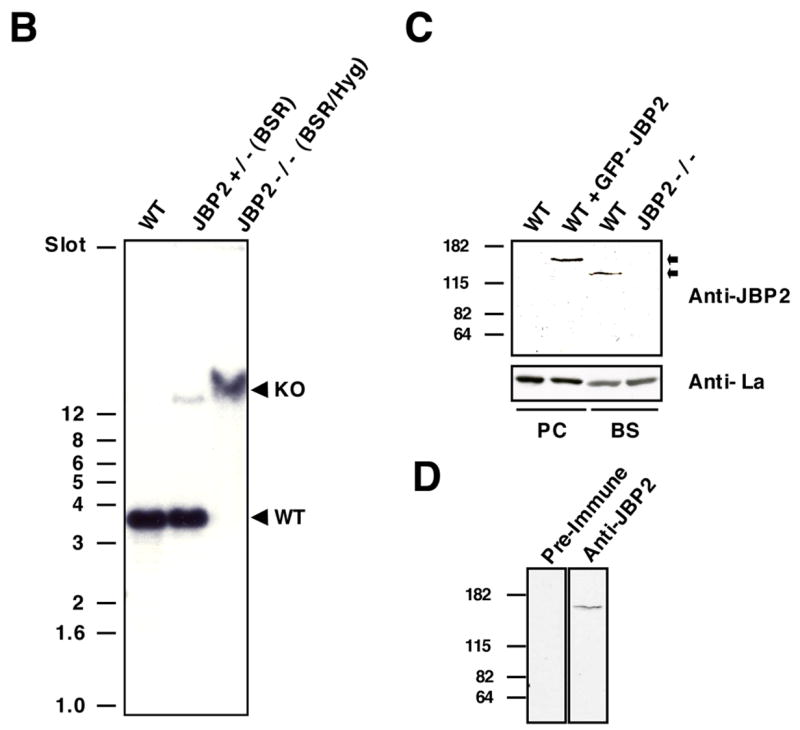
Synthesis of the modified thymine base, beta-d-glucosyl-hydroxymethyluracil or J, within telomeric DNA of Trypanosoma brucei correlates with the bloodstream form specific epigenetic silencing of telomeric variant surface glycoprotein genes involved in antigenic variation. In order to analyze the function of base J in the regulation of antigenic variation, we are characterizing the regulatory mechanism of J biosynthesis. We have recently proposed a model in which chromatin remodeling by a SWI2/SNF2-like protein (JBP2) regulates the developmental and de novo site-specific localization of J synthesis within bloodstream form trypanosome DNA. Consistent with this model, we now show that JBP2 (-/-) bloodstream form trypanosomes contain five-fold less base J and are unable to stimulate de novo J synthesis in newly generated telomeric arrays.
Figures




References
-
- Gommers-Ampt JH, Van Leeuwen F, de Beer AL, et al. beta-D-glucosyl-hydroxymethyluracil: a novel modified base present in the DNA of the parasitic protozoan T. brucei. Cell. 1993;75(6):1129–36. - PubMed
-
- Borst P, van Leeuwen F. beta-D-glucosyl-hydroxymethyluracil, a novel base in African trypanosomes and other Kinetoplastida. Molecular & Biochemical Parasitology. 1997;90(1):1–8. - PubMed
-
- van Leeuwen F, Taylor MC, Mondragon A, Moreau H, Gibson W, Kieft R, Borst P. beta-D-glucosyl-hydroxymethyluracil is a conserved DNA modification in kinetoplastid protozoans and is abundant in their telomeres. Proceedings of the National Academy of Sciences of the United States of America. 1998;95(5):2366–71. [see comments] - PMC - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Medical
