

Multiple cardiovascular defects caused by the absence of alternatively spliced segments of fibronectin
- PMID: 17706958
- PMCID: PMC2080666
- DOI: 10.1016/j.ydbio.2007.07.005
Multiple cardiovascular defects caused by the absence of alternatively spliced segments of fibronectin
Abstract
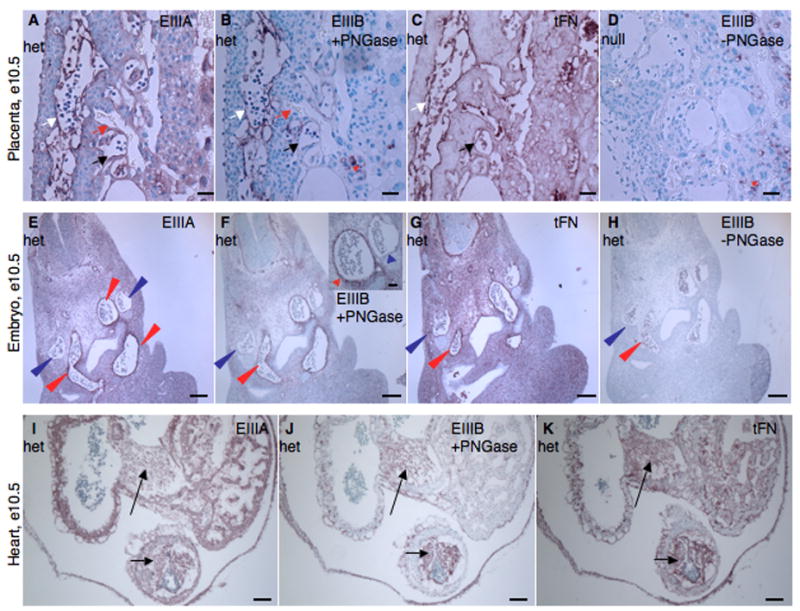
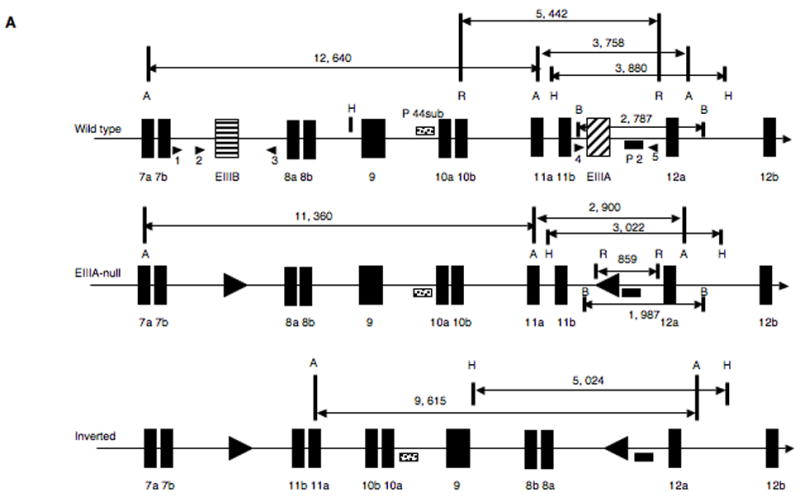
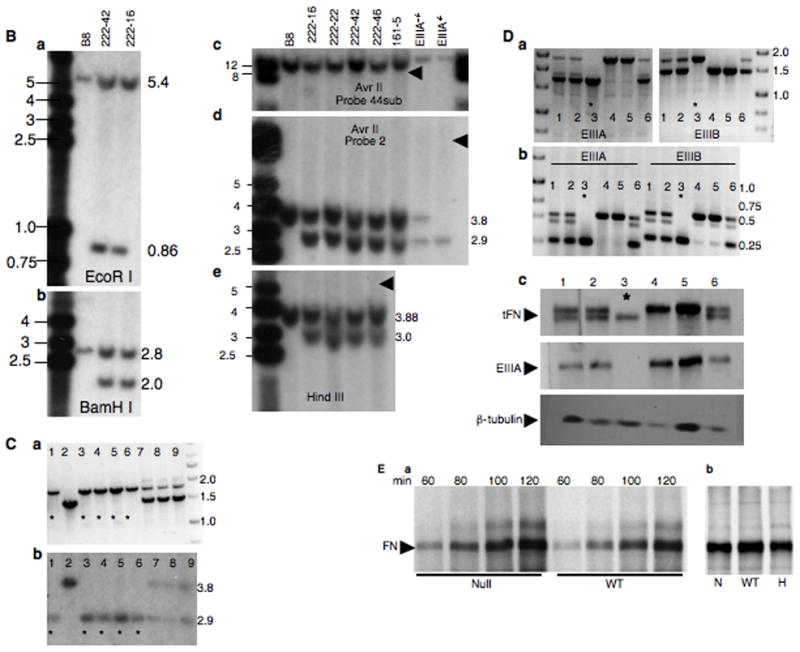
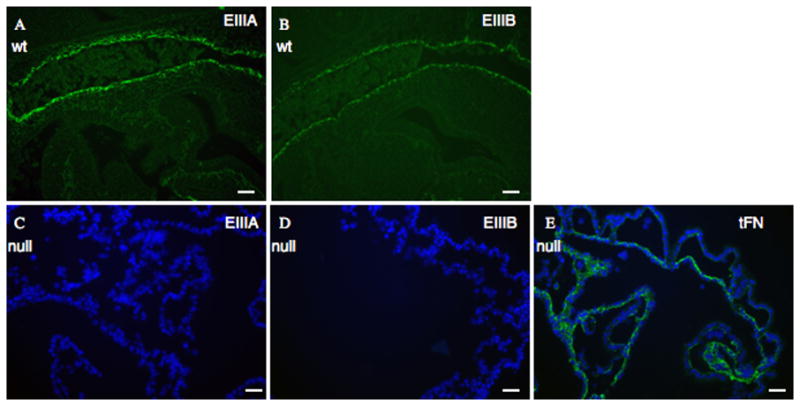
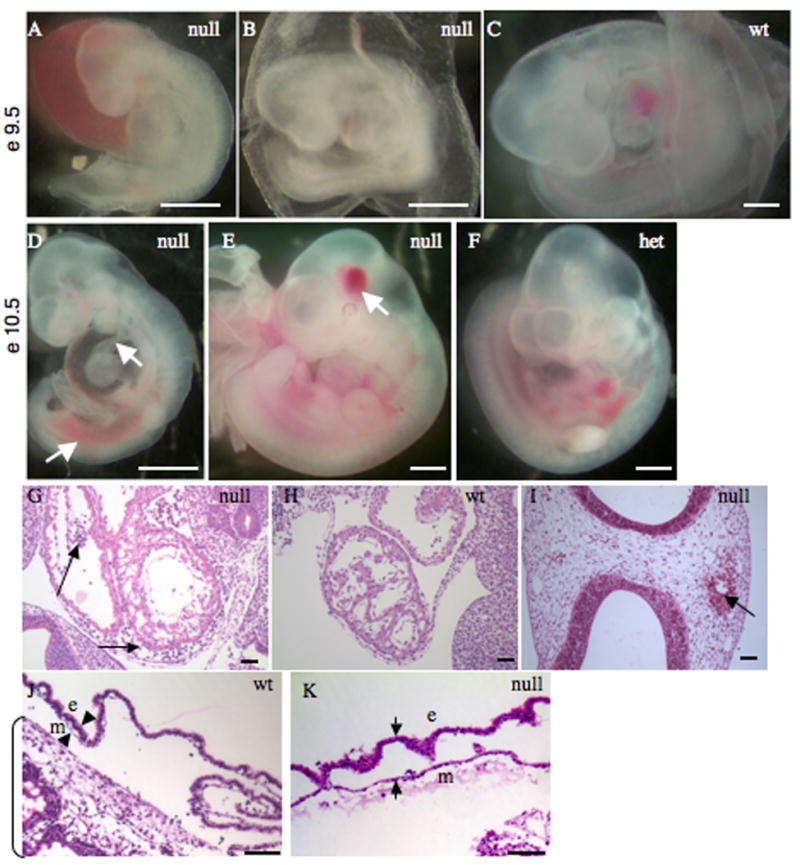
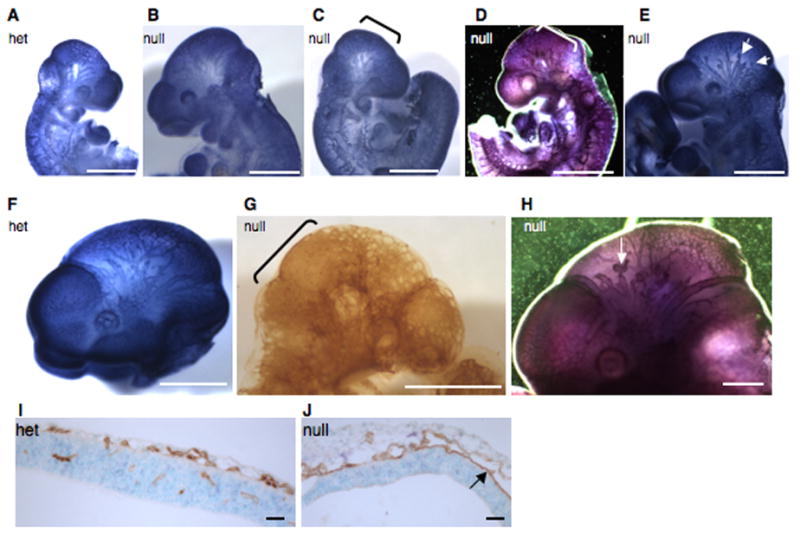
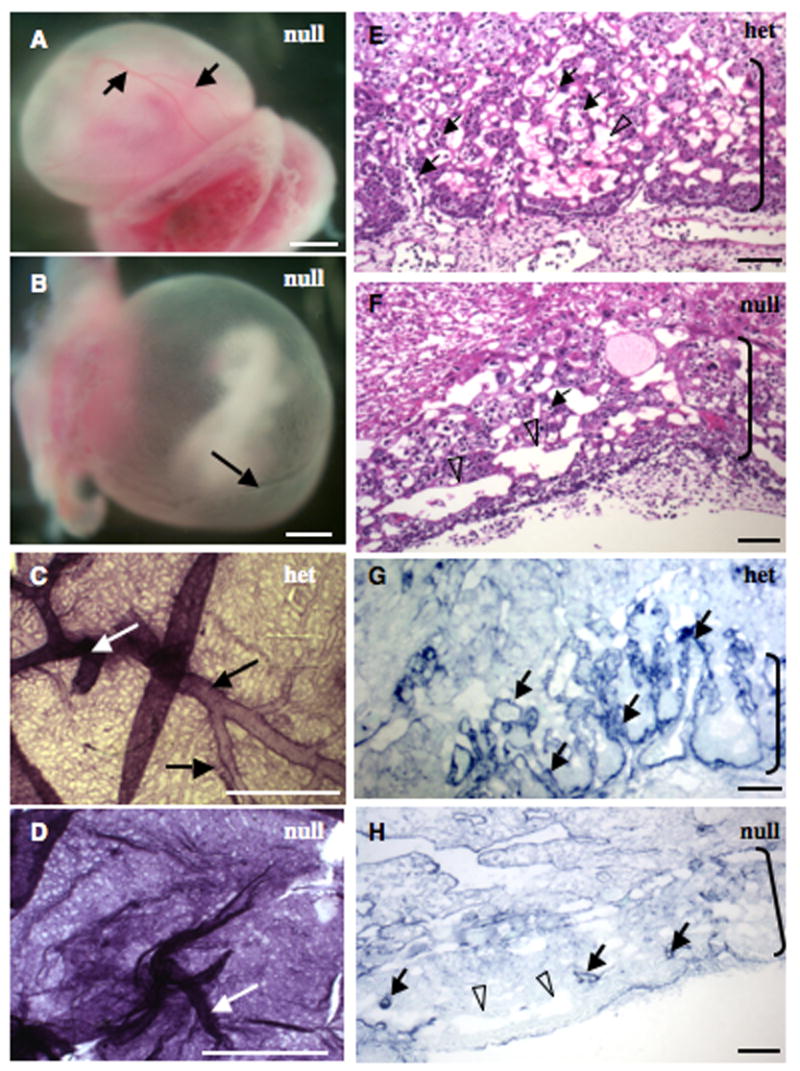
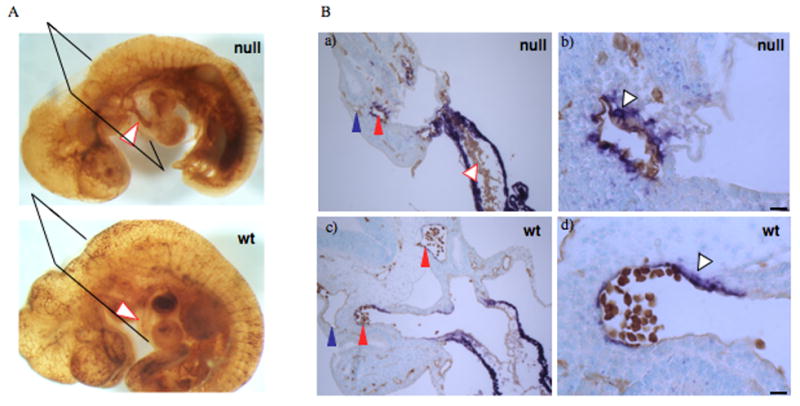
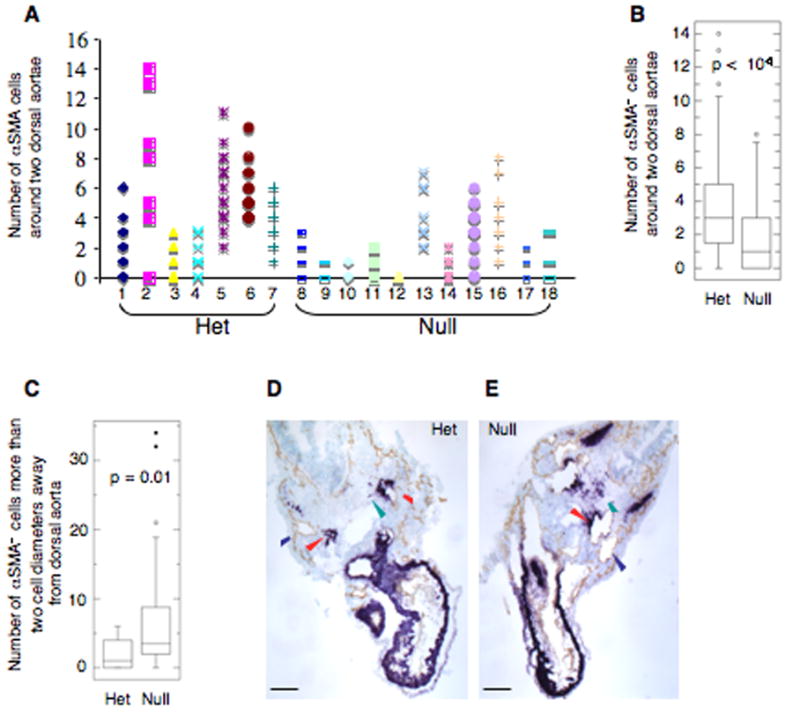
Alternatively spliced variants of fibronectin (FN) containing exons EIIIA and EIIIB are expressed around newly forming vessels in development and disease but are downregulated in mature vasculature. The sequences and patterns of expression of these splice variants are highly conserved among vertebrates, suggestive of their biological importance; however the functions of EIIIA and EIIIB-containing FNs are unknown. To understand the role(s) of these splice variants, we deleted both EIIIA and EIIIB exons from the FN gene and observed embryonic lethality with incomplete penetrance by embryonic day 10.5. Deletion of both EIIIA and EIIIB exons did not affect synthesis or cell surface deposition of FN, indicating that embryonic lethality was due specifically to the absence of EIIIA and EIIIB exons from FN. EIIIA/EIIIB double-null embryos displayed multiple embryonic cardiovascular defects, including vascular hemorrhage, failure of remodeling embryonic and yolk sac vasculature, defective placental angiogenesis and heart defects. In addition, we observed defects in coverage and association with dorsal aortae of alpha-smooth-muscle-actin-positive cells. Our studies indicate that the presence or absence of EIIIA and EIIIB exons alters the function of FN and demonstrate the requirement for these alternatively spliced exons in cardiovascular development.
Figures
References
-
- Anson-Cartwright L, Dawson K, Holmyard D, Fisher SJ, Lazzarini RA, Cross JC. The glial cells missing-1 protein is essential for branching morphogenesis in the chorioallantoic placenta. Nat Genet. 2000;25:311–4. - PubMed
-
- Aota S, Nagai T, Yamada KM. Characterization of regions of fibronectin besides the arginine-glycine-aspartic acid sequence required for adhesive function of the cell-binding domain using site-directed mutagenesis. J Biol Chem. 1991;266:15938–43. - PubMed
-
- Borsi L, Balza E, Bestagno M, Castellani P, Carnemolla B, Biro A, Leprini A, Sepulveda J, Burrone O, Neri D, et al. Selective targeting of tumoral vasculature: comparison of different formats of an antibody (L19) to the ED-B domain of fibronectin. Int J Cancer. 2002;102:75–85. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
Research Materials
Miscellaneous
