

Biphasic Hoxd gene expression in shark paired fins reveals an ancient origin of the distal limb domain
- PMID: 17710153
- PMCID: PMC1937022
- DOI: 10.1371/journal.pone.0000754
Biphasic Hoxd gene expression in shark paired fins reveals an ancient origin of the distal limb domain
Abstract
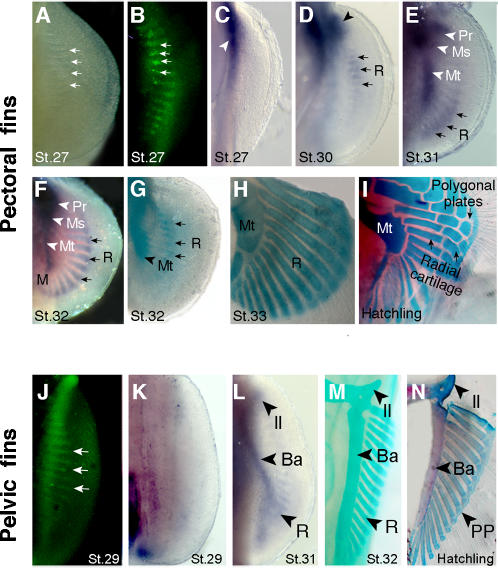
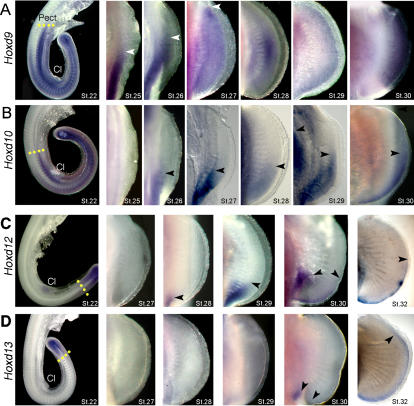
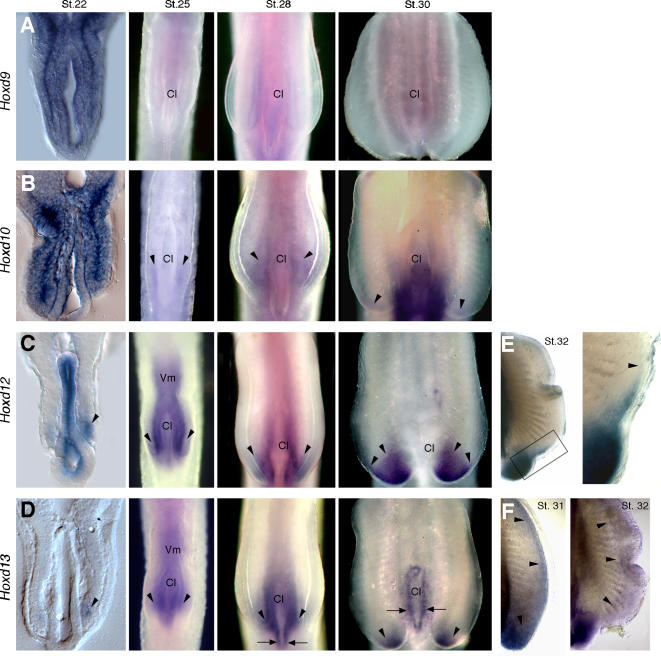
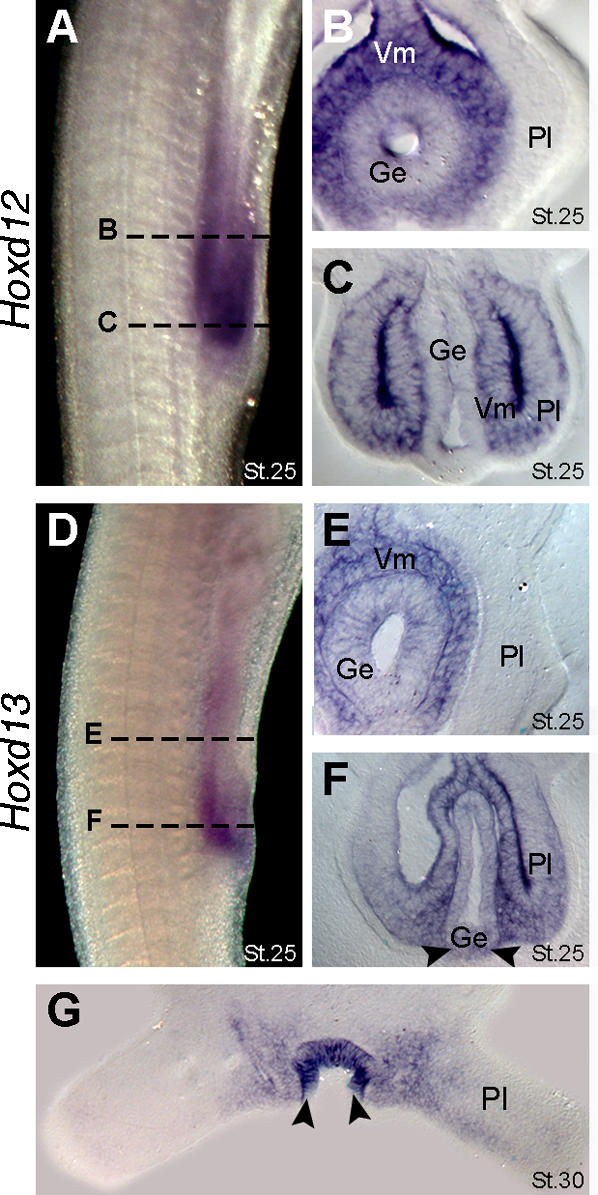
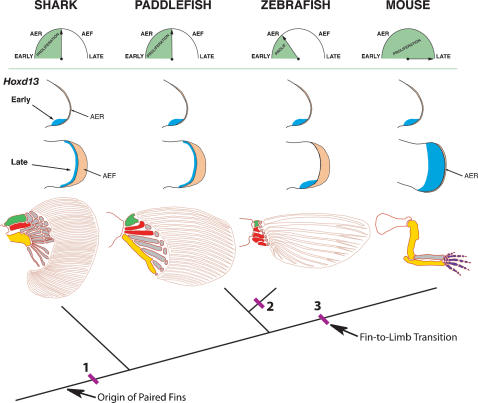
The evolutionary transition of fins to limbs involved development of a new suite of distal skeletal structures, the digits. During tetrapod limb development, genes at the 5' end of the HoxD cluster are expressed in two spatiotemporally distinct phases. In the first phase, Hoxd9-13 are activated sequentially and form nested domains along the anteroposterior axis of the limb. This initial phase patterns the limb from its proximal limit to the middle of the forearm. Later in development, a second wave of transcription results in 5' HoxD gene expression along the distal end of the limb bud, which regulates formation of digits. Studies of zebrafish fins showed that the second phase of Hox expression does not occur, leading to the idea that the origin of digits was driven by addition of the distal Hox expression domain in the earliest tetrapods. Here we test this hypothesis by investigating Hoxd gene expression during paired fin development in the shark Scyliorhinus canicula, a member of the most basal lineage of jawed vertebrates. We report that at early stages, 5'Hoxd genes are expressed in anteroposteriorly nested patterns, consistent with the initial wave of Hoxd transcription in teleost and tetrapod paired appendages. Unexpectedly, a second phase of expression occurs at later stages of shark fin development, in which Hoxd12 and Hoxd13 are re-expressed along the distal margin of the fin buds. This second phase is similar to that observed in tetrapod limbs. The results indicate that a second, distal phase of Hoxd gene expression is not uniquely associated with tetrapod digit development, but is more likely a plesiomorphic condition present the common ancestor of chondrichthyans and osteichthyans. We propose that a temporal extension, rather than de novo activation, of Hoxd expression in the distal part of the fin may have led to the evolution of digits.
Conflict of interest statement
Figures
References
-
- Coates MI. The evolution of paired fins. Theory in Biosciences. 2003;122:266–287.
-
- Shubin NH, Daeschler EB, Jenkins FA., Jr The pectoral fin of Tiktaalik roseae and the origin of the tetrapod limb. Nature. 2006;440:764–771. - PubMed
-
- Grandel H. Approaches to a comparison of fin and limb structure and development. Theory Biosci. 2003;122:288–301.
-
- Zhang XG, Hou XG. Evidence for a single median fin-fold and tail in the Lower Cambrian vertebrate, Haikouichthys ercaicunensis. J Evol Biol. 2004;17:1162–1166. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
