

Combining phylogeography with distribution modeling: multiple Pleistocene range expansions in a parthenogenetic gecko from the Australian arid zone
- PMID: 17712408
- PMCID: PMC1942116
- DOI: 10.1371/journal.pone.0000760
Combining phylogeography with distribution modeling: multiple Pleistocene range expansions in a parthenogenetic gecko from the Australian arid zone
Abstract
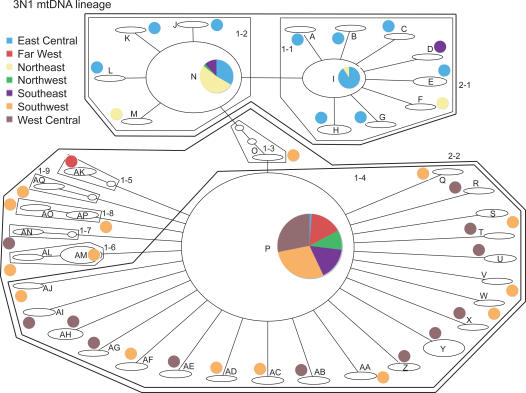
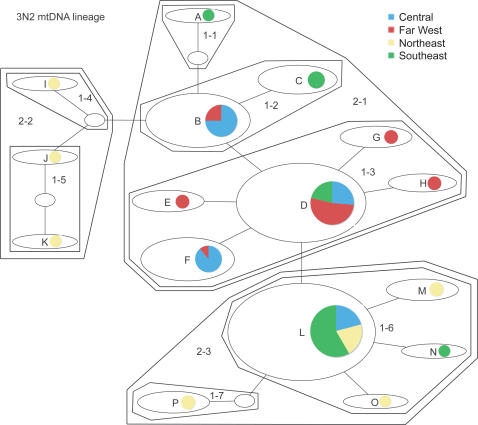
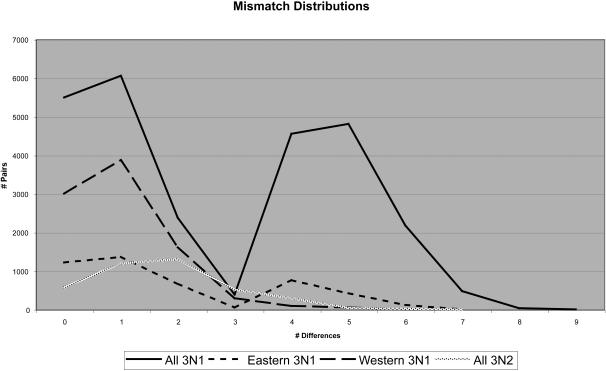
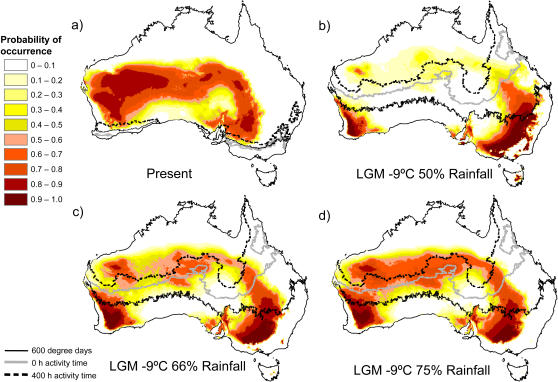
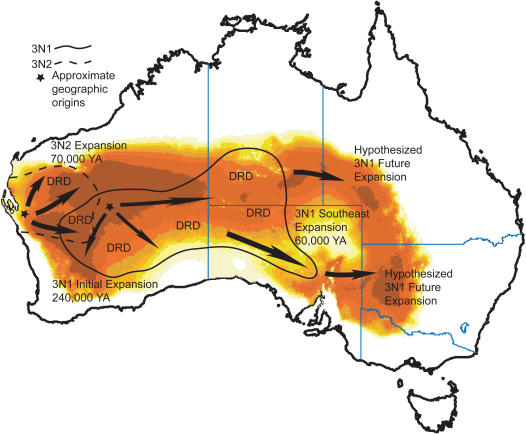
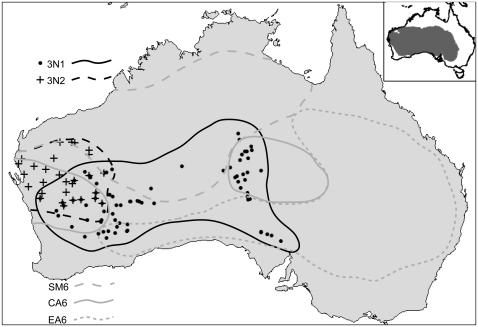
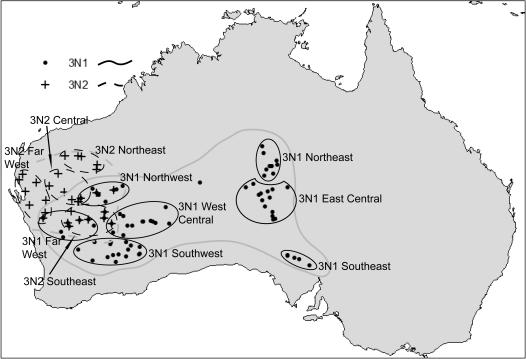
Phylogenetic and geographic evidence suggest that many parthenogenetic organisms have evolved recently and have spread rapidly. These patterns play a critical role in our understanding of the relative merits of sexual versus asexual reproductive modes, yet their interpretation is often hampered by a lack of detail. Here we present a detailed phylogeographic study of a vertebrate parthenogen, the Australian gecko Heteronotia binoei, in combination with statistical and biophysical modeling of its distribution during the last glacial maximum. Parthenogenetic H. binoei occur in the Australian arid zone and have the widest range of any known vertebrate parthenogen. They are broadly sympatric with their sexual counterparts, from which they arose via hybridization. We have applied nested clade phylogeographic, effective migration, and mismatch distribution analyses to mitochondrial DNA (mtDNA) sequences obtained for 319 individuals sampled throughout the known geographic ranges of two parthenogenetic mitochondrial lineages. These analyses provide strong evidence for past range expansion events from west to east across the arid zone, and for continuing eastward range expansion. Parthenogen formation and range expansion events date to the late Pleistocene, with one lineage expanding from the northwest of its present range around 240,000 years ago and the second lineage expanding from the far west around 70,000 years ago. Statistical and biophysical distribution models support these inferences of recent range expansion, with suitable climatic conditions during the last glacial maximum most likely limited to parts of the arid zone north and west of much of the current ranges of these lineages. Combination of phylogeographic analyses and distribution modeling allowed considerably stronger inferences of the history of this complex than either would in isolation, illustrating the power of combining complementary analytical approaches.
Conflict of interest statement
Figures
Similar articles
-
Phylogeography of sexual Heteronotia binoei (Gekkonidae) in the Australian arid zone: climatic cycling and repetitive hybridization.Mol Ecol. 2005 Aug;14(9):2755-72. doi: 10.1111/j.1365-294X.2005.02627.x. Mol Ecol. 2005. PMID: 16029476
-
The evolution of sexual and parthenogenetic Warramaba: a window onto Plio-Pleistocene diversification processes in an arid biome.Mol Ecol. 2008 Dec;17(24):5257-75. doi: 10.1111/j.1365-294X.2008.03991.x. Mol Ecol. 2008. PMID: 19120998
-
Not always young: The first vertebrate ancient origin of true parthenogenesis found in an Amazon leaf litter lizard with evidence of mitochondrial haplotypes surfing on the wave of a range expansion.Mol Phylogenet Evol. 2019 Jun;135:105-122. doi: 10.1016/j.ympev.2019.01.023. Epub 2019 Feb 4. Mol Phylogenet Evol. 2019. PMID: 30731120
-
Birth of a biome: insights into the assembly and maintenance of the Australian arid zone biota.Mol Ecol. 2008 Oct;17(20):4398-417. doi: 10.1111/j.1365-294X.2008.03899.x. Epub 2008 Aug 27. Mol Ecol. 2008. PMID: 18761619 Review.
-
Impacts of Quaternary glaciation, geological history and geography on animal species history in continental East Asia: A phylogeographic review.Mol Ecol. 2023 Aug;32(16):4497-4514. doi: 10.1111/mec.17053. Epub 2023 Jun 18. Mol Ecol. 2023. PMID: 37332105 Review.
Cited by
-
Coherent and incoherent inference in phylogeography and human evolution.Proc Natl Acad Sci U S A. 2010 Apr 6;107(14):6376-81. doi: 10.1073/pnas.0910647107. Epub 2010 Mar 22. Proc Natl Acad Sci U S A. 2010. PMID: 20308555 Free PMC article.
-
Multi-Species Phylogeography of Arid-Zone Sminthopsinae (Marsupialia: Dasyuridae) Reveals Evidence of Refugia and Population Expansion in Response to Quaternary Change.Genes (Basel). 2020 Aug 20;11(9):963. doi: 10.3390/genes11090963. Genes (Basel). 2020. PMID: 32825338 Free PMC article.
-
Nested clade analysis: an extensively validated method for strong phylogeographic inference.Mol Ecol. 2008 Apr;17(8):1877-80. doi: 10.1111/j.1365-294X.2008.03731.x. Epub 2008 Mar 10. Mol Ecol. 2008. PMID: 18346121 Free PMC article. No abstract available.
-
Inference of population history by coupling exploratory and model-driven phylogeographic analyses.Int J Mol Sci. 2010 Mar 24;11(4):1190-227. doi: 10.3390/ijms11041190. Int J Mol Sci. 2010. PMID: 20480016 Free PMC article. Review.
-
Integrating statistical and mechanistic approaches with biotic and environmental variables improves model predictions of the impact of climate and land-use changes on future mosquito-vector abundance, diversity and distributions in Australia.Parasit Vectors. 2020 Sep 23;13(1):484. doi: 10.1186/s13071-020-04360-3. Parasit Vectors. 2020. PMID: 32967711 Free PMC article. Review.
References
-
- Dawley RM, Bogart JP. Albany): (New York State Museum Press; 1989.
-
- White MJD. Modes of Speciation. San Francisco): (W. H. Freeman; 1978.
-
- Vrijenhoek RC, Dawley RM, Cole CC, Bogart JP. In: Evolution and Ecology of Unisexual Vertebrates, Dawley RM, Bogart JP, editors. Albany): (New York State Museum; 1989. pp. 19–23.
-
- Bell G. The Masterpiece of Nature: the Evolution and Genetics of Sexuality. Berkeley): (University of California Press; 1982.
-
- Butlin R. Nature Reviews Genetics. 2002;3:311–317. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Miscellaneous
