

The amphioxus SoxB family: implications for the evolution of vertebrate placodes
- PMID: 17713598
- PMCID: PMC1950271
- DOI: 10.7150/ijbs.3.356
The amphioxus SoxB family: implications for the evolution of vertebrate placodes
Abstract
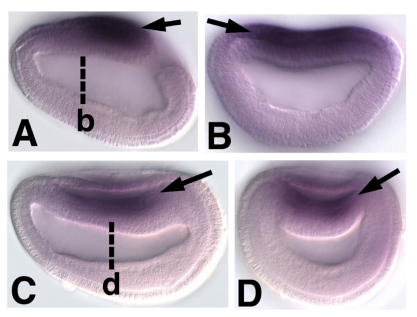
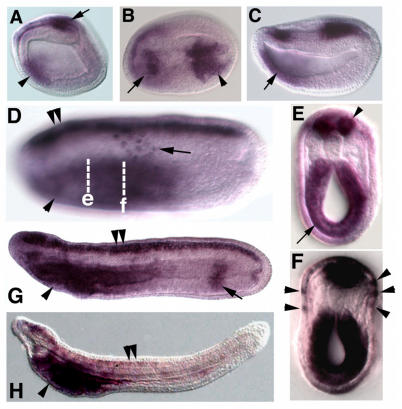
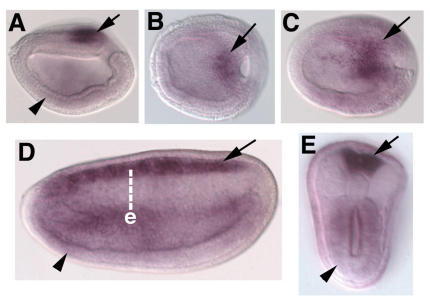
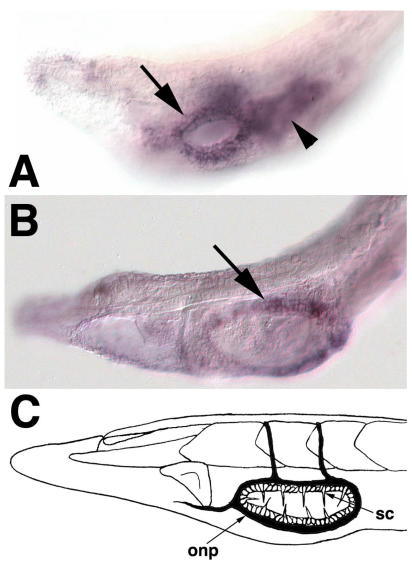
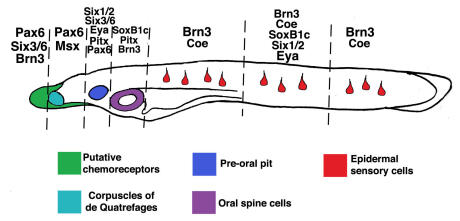
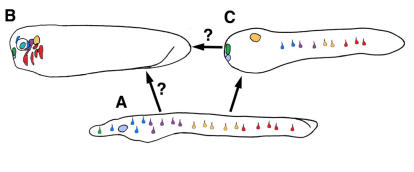
Cranial placodes are regions of thickened ectoderm that give rise to sense organs and ganglia in the vertebrate head. Homologous structures are proposed to exist in urochordates, but have not been found in cephalochordates, suggesting the first chordates lacked placodes. SoxB genes are expressed in discrete subsets of vertebrate placodes. To investigate how placodes arose and diversified in the vertebrate lineage we isolated the complete set of SoxB genes from amphioxus and analyzed their expression in embryos and larvae. We find that while amphioxus possesses a single SoxB2 gene, it has three SoxB1 paralogs. Like vertebrate SoxB1 genes, one of these paralogs is expressed in non-neural ectoderm destined to give rise to sensory cells. When considered in the context of other amphioxus placode marker orthologs, amphioxus SoxB1 expression suggests a diversity of sensory cell types utilizing distinct placode-type gene programs was present in the first chordates. Our data supports a model for placode evolution and diversification whereby the full complement of vertebrate placodes evolved by serial recruitment of distinct sensory cell specification programs to anterior pre-placodal ectoderm.
Keywords: Evolution; amphioxus; chordates; development; placodes; vertebrates.
Conflict of interest statement
CONFLICT OF INTEREST: The authors have declared that no conflict of interest exists.
Figures

Similar articles
-
The evolutionary history of vertebrate cranial placodes II. Evolution of ectodermal patterning.Dev Biol. 2014 May 1;389(1):98-119. doi: 10.1016/j.ydbio.2014.01.019. Epub 2014 Feb 1. Dev Biol. 2014. PMID: 24491817 Review.
-
Evolutionary origins of vertebrate placodes: insights from developmental studies and from comparisons with other deuterostomes.J Exp Zool B Mol Dev Evol. 2005 Jul 15;304(4):347-99. doi: 10.1002/jez.b.21055. J Exp Zool B Mol Dev Evol. 2005. PMID: 16003766 Review.
-
From so simple a beginning - what amphioxus can teach us about placode evolution.Int J Dev Biol. 2017;61(10-11-12):633-648. doi: 10.1387/ijdb.170127gs. Int J Dev Biol. 2017. PMID: 29319112 Review.
-
Evolutionary conservation of the presumptive neural plate markers AmphiSox1/2/3 and AmphiNeurogenin in the invertebrate chordate amphioxus.Dev Biol. 2000 Oct 1;226(1):18-33. doi: 10.1006/dbio.2000.9810. Dev Biol. 2000. PMID: 10993671
-
Conservation and elaboration of Hox gene regulation during evolution of the vertebrate head.Nature. 2000 Dec 14;408(6814):854-7. doi: 10.1038/35048570. Nature. 2000. PMID: 11130723
Cited by
-
Analyses of Sox-B and Sox-E Family Genes in the Cephalopod Sepia officinalis: Revealing the Conserved and the Unusual.PLoS One. 2016 Jun 22;11(6):e0157821. doi: 10.1371/journal.pone.0157821. eCollection 2016. PLoS One. 2016. PMID: 27331398 Free PMC article.
-
Genome-wide analysis of the sox family in the calcareous sponge Sycon ciliatum: multiple genes with unique expression patterns.Evodevo. 2012 Jul 23;3(1):14. doi: 10.1186/2041-9139-3-14. Evodevo. 2012. PMID: 22824100 Free PMC article.
-
Evolution of non-coding regulatory sequences involved in the developmental process: reflection of differential employment of paralogous genes as highlighted by Sox2 and group B1 Sox genes.Proc Jpn Acad Ser B Phys Biol Sci. 2009;85(2):55-68. doi: 10.2183/pjab.85.55. Proc Jpn Acad Ser B Phys Biol Sci. 2009. PMID: 19212098 Free PMC article. Review.
-
Gene Regulatory Networks of Epidermal and Neural Fate Choice in a Chordate.Mol Biol Evol. 2022 Apr 11;39(4):msac055. doi: 10.1093/molbev/msac055. Mol Biol Evol. 2022. PMID: 35276009 Free PMC article.
-
The evolutionary origins of the vertebrate olfactory system.Open Biol. 2020 Dec;10(12):200330. doi: 10.1098/rsob.200330. Epub 2020 Dec 23. Open Biol. 2020. PMID: 33352063 Free PMC article. Review.
References
-
- Bassham S, Postlethwait JH. The evolutionary history of placodes: a molecular genetic investigation of the larvacean urochordate Oikopleura dioica. Development. 2005;132:4259–72. - PubMed
-
- Mazet F, Hutt JA, Milloz J, Millard J, Graham A, Shimeld SM. Molecular evidence from Ciona intestinalis for the evolutionary origin of vertebrate sensory placodes. Dev Biol. 2005;282:494–508. - PubMed
-
- Delsuc F, Brinkmann H, Chourrout D, Philippe H. Tunicates and not cephalochordates are the closest living relatives of vertebrates. Nature. 2006;439:965–8. - PubMed
-
- Blair JE, Hedges SB. Molecular phylogeny and divergence times of deuterostome animals. Mol Biol Evol. 2005;22:2275–84. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
